")
Back to Journals » Clinical, Cosmetic and Investigational Dermatology » Volume 15
Anti-Glycation and Anti-Aging Properties of Resveratrol Derivatives in the in-vitro 3D Models of Human Skin
Authors Markiewicz E , Jerome J, Mammone T, Idowu OC
Received 3 March 2022
Accepted for publication 3 May 2022
Published 19 May 2022 Volume 2022:15 Pages 911—927
DOI https://doi.org/10.2147/CCID.S364538
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Jeffrey Weinberg
Ewa Markiewicz,1 Jaimie Jerome,2 Thomas Mammone,2 Olusola C Idowu1
1Hexis Lab Limited, The Catalyst, Newcastle Helix, Newcastle upon Tyne, UK; 2Estee Lauder Research Laboratories, Melville, NY, USA
Correspondence: Thomas Mammone, Estee Lauder Research Laboratories, Melville, NY, USA, Tel +1 1 877 311 3883, Email [email protected] Olusola C Idowu, HexisLab Limited, The Catalyst, Newcastle Helix, Newcastle upon Tyne, NE4 5TG, UK, Tel +44 1394 825487, Email [email protected]
Purpose: Human skin undergoes modifications affecting its structural properties and barrier functions involved in protection against age-related damage. Glycation is a non-enzymatic reaction between macromolecules and sugars causing alterations to the elastic fibers and premature aging of the skin. Glycation can be prevented by a range of bioactive molecules; however, at present only a few of them are validated for inclusion in cosmetic products. There is also a demand for reproducible in-vitro assays demonstrating the anti-aging effect of compounds on the skin. This study aimed to define the potential targets for screening and validation of anti-glycation activity of novel cosmetic candidates from natural products and to provide a plausible mechanism for their anti-aging potential based on 3D skin models.
Methods: Dermal fibroblasts and 3D skin models were treated with glycation agent and topical applications of Resveratrol derivatives. The samples were analyzed for advanced glycation end products (AGEs) alongside an organization of elastic fibers and expression of proliferative, senescence, and oxidative stress markers by autofluorescence, immunocytochemistry and quantitative assays.
Results: Accumulation of AGEs in the 3D skin model is associated with reduced stratification of the epidermis and re-organization of the collagen in the upper, cell-dense layer of the dermis. Treatment of dermal fibroblasts with Resveratrol, OxyResveratrol, Piceatannol, and Triacetyl Resveratrol ameliorates the effects of glycation consistent with cellular aging. Subsequent topical application of the compounds in skin models results in a reduction in glycation-induced AGEs, an increase in collagen expression and a stratification of the epidermis.
Conclusion: Glycation could result in age-related alterations in the structural and cellular organizations of the superficial layers of the skin, which can be restored by Resveratrol derivatives, pointing to their promising capacities as bioactive ingredients in cosmetic products. Insight into the potential parameters affected by skin glycation could also serve as a reference for screening the bioactive molecules for cosmetic purposes.
Keywords: glycation, skin aging, collagen, fibroblast, resveratrol
Introduction
Glycation is a physiological, non-enzymatic reaction between free reducing sugars and proteins, DNA and lipids that can damage the tissue.1 The process is typically initiated by the reactive carbonyl groups of sugars, resulting in intermediate Amadori products that are modified further by oxidation, dehydration, polymerization and cross-linking leading to the formation and advanced glycation end products (AGEs). Glycation frequently involves the modification of the arginine residues of the protein with metabolic factors, such as Glyoxal, Methylglyoxal and 3-Deoxyglucosone.2 There is also a close biological association between the formation of AGEs-modified macromolecules and glycemic index-linked to tissue damage in early and advanced metabolic syndromes that were recorded in clinical trials.3,4 In addition, glycated macromolecules can be formed during oxidative stress due to hyperglycemia and plasma levels of circulating AGEs often correspond to glycemic index control.5
Accumulation of the glycated macromolecules, predominantly proteins, has been shown to impair organ function and is regarded as a plausible hallmark of tissue damage linked to aging and age-related diseases.6 There is also extensive experimental and theoretical evidence pointing at the accumulation of AGEs in the tissues of aged animal models, frequently linked to metabolism-mediated life span.7,8 The AGEs that have been extensively characterized in this context include pentosidine, N-(carboxyethyl)-lysine (CEL) and N-(Carboxymethyl)-lysine (CML), which accumulate in several aged tissues including the eye, heart and arteries.9,10
There are several theoretical explanations and empirically supported mechanisms for the glycation-induced damage specifically affecting aged tissues. The studies point at glycation as a cause of the damage affecting mitochondrial proteins involved in fatty acid b-oxidation and tricarboxylic acid (TCA) cycle leading to a decline in capacity for energy production.11 Post-translation modification of proteins by glycation could also interfere with the process of angiogenesis in aging and age-related disorders.12 Finally, the formation of AGEs and the macromolecular damage are facilitated by an increase in oxidative stress and controlled by feedback involving reactive oxygen and nitrogen species (ROS and NOS) and the antioxidant capacities of the cells.13
Events of glycation have also been extensively studied in the skin. As a large, stratified organ, the skin is composed of two main layers with defined structure and function: the epidermis, the outermost layer formed predominantly by keratinocytes and providing a protective barrier, and the dermis, formed predominantly by dermal fibroblasts, elastic fibers and collagen contributing to the mechanical strength and overall skin elasticity.14 Effects of glycation on the skin include impairment in the mechanical propensity, wound healing and angiogenesis, with the accumulation of AGEs directly linked to the diet.15 Critical effects of glycation on skin aging are indicated by the impact of AGEs on the physical and biological properties of the collagen and elastic fibers affecting the architecture of the dermis and extracellular matrix (ECM) remodeling.16 The major mechanisms contributing to the damage involve the accumulation of AGEs within the ECM and chemical reactions typically leading to a significant decrease in the connective tissue elasticity and increased stiffness. Such changes are frequently accompanied by activation of the inflammatory and oxidative stress pathways and are further exasperated by low expression or absence of the enzymes removing the glycated products. In aged tissues and the skin, the most prevalent AGEs modifications involve CML and pentosidine, which accumulate with time and can permanently alter the structure of collagen and elastic fibers.17 Glycation of collagen coincides with a decrease in its turnover, intermolecular cross-linking, impaired remodeling and interactions with the cell.18,19 AGEs modifications are also detected in the epidermis, where they could be potentially linked to changes in the epidermal structure and homeostasis.20
Several natural bioactive compounds have been recently identified as promising candidates with the ability to inhibit glycation and glycation-mediated skin aging. Compared with synthetic molecules, the small molecules from natural sources are generally considered safe for both consumption and topical applications.21 Such ingredients have also added protective beneficial effects stemming from their biological functions, such as antioxidants, which can potentiate the anti-glycation activity in the cell. The effect of the compounds is evident in the inhibition of AGEs formation and trapping of reactive dicarbonyl species, which are frequently related to the precise modification and chemical structure of the compound.22–24 For some ingredients, the anti-glycation capacities appear similar or even stronger than for synthetic compounds such as aminoguanidine originally identified in the in vivo studies.25 Therefore, the naturally occurring small bioactive molecules can be evaluated further and incorporated into cosmetic formulations with skincare benefits.
In this study, we investigated the effect of glycation on dermal-epidermal compartments and cellular renewal in an in-vitro 3D skin model. We found that AGEs-related damage to the collagen and elastic fibers is closely associated with the alterations in the fibroblast-dense upper layer of the dermis, which interacts directly with the epidermis, leading to a decrease in the epidermal thickness and stratification. Treatment of cultured dermal fibroblasts with Methylglyoxal induces senescent phenotypes of the cells, such characteristics can be prevented by the presence of Resveratrol and its derivatives, OxyResveratrol, Piceatannol and Triacetyl Resveratrol in growth media. Subsequent topical applications of the aqueous solutions of the compounds in skin models ameliorate the effects of glycation, including inhibition of AGEs formation, increase in fibroblasts and collagen density, and improved epidermal stratification. These data provide further insight into the anti-aging propensity of Resveratrol and its derivatives and inform the potential novel applications for cosmetic formulations based on the anti-glycation activity.
Materials and Methods
Full-Thickness Skin Models and Dermal Fibroblasts Cultures
Full-thickness skin models, 1 cm in diameter (EpiDermFT) and Assay/Maintenance medium (EFT-400) were purchased from MatTek, In Vitro Life Science Laboratories. The models were equilibrated for 24 hours in 2.5 mL maintenance media in 6 well plates at the air–liquid interface in 5% CO2, 95% humidified incubator at 37°C. Carnosine (C9625-5G, Sigma), Resveratrol (34092-100MG, Sigma), Oxyresveratrol (91211-100MG, Sigma), Piceatannol (PO453-5MG, Sigma), and Triacetyl Resveratrol (SML0032-10MG, Sigma) were solubilized in filter sterile DMSO and diluted in phosphate-buffered saline (PBS) to 0.5% aqueous solution. After 24 hours, the maintenance media was replaced with fresh and the models were pre-treated with compound solutions, applied topically, 50 mL per model, in triplicates. Control models were treated with PBS only, in triplicates. The next day (day 0) the models were treated with Methylglyoxal (MGO, Sigma), added directly to the growth media (bottom chamber) at a concentration of 500 mM in 5 mL media. On days 2, 4, and 6 the models were treated with fresh maintenance media containing MGO and the topically applied compounds and fixed for analysis on day 7. Control models were treated with MGO only, in triplicates.
Human dermal fibroblasts (HDFs) were purchased from ThermoFisher and the cultures established in Dulbecco’s Modified Eagle’s Medium (DMEM) with 10% Fetal Bovine Serum (FBS, ThermoFisher) in 5% CO2, 95% humidified incubator at 37°C and passaged using Trypsin-EDTA buffer (Gibco). The cells were pre-treated with 0.01% solutions of Carnosine, Resveratrol, Oxyresveratrol, Piceatannol and Triacetyl Resveratrol in growth media for 24 hours, in triplicate samples for each compound. The next day, 500 mM MGO was added to the media and the cultures were maintained for 72 hours before analysis. Control samples were the cells -MGO/-compounds and +MGO/-compounds, in triplicates.
Measurements of MGO-Induced AGEs by Auto-Fluorescence
For the measurements of MGO-induced AGEs accumulation, EpiDermFT skin models or HDF cultures were washed in phosphate-buffered saline (PBS) and the 6-well plates were transferred to a microplate reader (SpectraMax iD5 Microplate Reader, Molecular Devices). The intensities of the fluorescence signals were recorded at Ex 370 nm/Em 440 nm and Ex 335 nm/Em 385 nm on days 0, 1, 4 and 7 and the relative fluorescence signal (RFI) was normalized to control.
For the visualization of the auto-fluorescence, the samples were observed under a fluorescence microscope (Leica DM IL LED, DAPI channel) and photographed using CCD DFC3000G Camera and LAS X 3.6.0.20104 software (Leica Microsystems).
Histochemical Processing of the Samples
The skin samples were fixed in 4% formaldehyde/PBS overnight at 4°C and processed as follows: 30% ethanol, 30 min.; 50% ethanol 30 min.; 70% ethanol 30 min.; 80% ethanol 30 min.; 90% ethanol 30 min.; 95% ethanol 30 min.; 100% ethanol 30 min.; Histoclear 30 min.; 50%:50% solution of Histoclear and molten paraffin wax, 30 min. at 60°C.; 100% molten paraffin wax, 60 min. at 60°C and wax-embedded in the embedding molds. The samples were cut using a microtome (HM340E Electronic Rotary Microtome, Fisher Scientific) at the 5 µm specimen thickness, adhered to glass slides (SuperFrost Plus, Thermo Fisher Scientific) in a 50 °C water bath and left to dry overnight. For the analysis, the sections were stained with hematoxylin and eosin (H&E) as follows: Histoclear 5 min.; 100% ethanol 2 min.; 95% ethanol 1 min.; 70% ethanol 1 min., water 1 min.; Haematoxylin 5 min.; water 30 sec.; alkaline alcohol 30 sec.; 70% ethanol 30 sec.; 95% ethanol 30 sec.; Eosin 30 sec.; 95% ethanol 30 sec.; 100% ethanol 30 sec; 2 × 5 min Histoclear.
For the staining of connective tissue, Masson’s trichrome stain kit (Sigma-Aldrich) was applied according to the manufacturer’s protocol. The slides were mounted in the Entellan mounting medium (Sigma).
The HDF cultures were fixed in 4% formaldehyde/PBS and stained for 15 minutes with 0.5% crystal violet diluted in 10% methanol and washed 3x H2O. All images were captured using Leica DMi1 Inverted Light Microscope with lens magnification 10X and 20X.
Immunostaining and Fluorescence Imaging
The skin sections samples were de-paraffinized and antigen retrieval was performed as follows: 2 × 5 min. Xylene; 2 × 5 min. 100% ethanol; 2 × 5 min. 70% ethanol; 2 × 5 min. water; Citrate buffer pH6, 30 min. at 95°C; 2 × 5 min. water; 5 min PBS (ThermoFisher). HDF cultures growing on glass coverslips were fixed in 4% formaldehyde/PBS for 15 minutes at RT, permeabilized with 0.5% TRITON X-100 for 5 minutes and washed 3 × PBS. For immunostaining, the slides with skin sections and the coverslips with HDF were incubated with primary antibodies diluted in the blocking buffer (PBS + 1% FBS) at 4°C overnight; washed 30 × 10 min. in PBS; incubated with secondary antibodies diluted in the blocking buffer (PBS+1% FBS) at room temp. 1 hr; washed 30 × 10 min. in PBS and mounted on the ProLong Gold Antifade Mountant with DAPI (ThermoFisher).
The primary antibodies were anti-CML (CML26, Abcam, 1:500), anti-Ki67 (D3B5, Cell Signaling, 1:500) and anti-p16 INK4a (E6N8P, Cell Signaling, 1:500). Secondary antibodies were TRITC anti-mouse and TRITC anti-rabbit (Jackson ImmunoResearch; 1:1000). The fluorescent images, including eosin, were captured using Leica DM IL LED and photographed using CCD DFC3000G Camera and LAS X 3.6.0.20104 software (Leica Microsystems) with lens magnification 10x and 20x. All figures were assembled using PowerPoint and Image J.
Biometric Measurements and Statistical Analysis
Biometric data were collected manually based on the Hematoxylin and Eosin (H&E), DAPI- and immuno-stained images of the 5 µm skin sections using ImageJ software (ImageJ 1.36b, National Institute of Health, USA). From the fluorescent and H&E-stained images, the initial measurements of the model sample length were performed on three consecutive images by tracing them alongside a straight line. The collected measurements of length units were then converted to microns, based on the 1380 pixels corresponding to the ImageJ 9 length units: 9 length units = 440 µm for fluorescent images (40x lens magnification) and 2048 pixels corresponding to the ImageJ 21 length units: 21 length units = 650 µm for H&E images (20x lens magnification). For densities of the dermal and epidermal cells, the cells were counted on each image and the collected numbers were recalculated per 0.1 mm2 of the area. For the epidermal thickness, the epidermis was traced manually, the measurements collected as Image J length units, and the obtained values converted to µm. For quantitative analysis of granular cellular populations, the cells were selected based on their nuclear shapes/orientation and localization in the epidermis. The respective numbers for each population were collected from each image and re-calculated per 1 mm of the sample. For densitometry analysis of CML expression and eosin density, the intensities of immunofluorescent signals were measured in a defined area on three consecutive images for each treatment using ImageJ software and adjusted to control sample values equal to 1. For calculations of cell densities and Ki67/p16INK4a positive cells, the cells were quantified based on manual counting and corresponding signal intensity in the defined area, with each population of antigen-positive cells represented as a percentage of total cells. The graphs represent the mean ± standard deviation, with statistically significant outputs * p < 0.05, ** p < 0.05, and ***p < 0.005 at 2 tailed paired t-test distribution.
Intracellular ROS Production Assay
For the measurements of intracellular ROS production, HDFs were seeded onto 96 well plates at a density of 2 × 103 per well and analyzed 24 hours post-treatment with the compounds. The medium was removed and replaced with PBS, and the cells were incubated for 30 minutes with a 10 mM 2ʹ7’-dichlorofluorescein diacetate (DCF-DA) probe (Abcam). Conversion into fluorescent DCF was induced by further incubation of the cells in DMEM/10% FBS in a CO2 incubator for 15 minutes. The cells were fixed with 4% formaldehyde/PBS, washed with PBS, and relative fluorescence intensity was recorded in a microplate reader (SpectraMax iD5 Microplate Reader, Molecular Devices) at Ex/Em 485/535 nm.
Results
Glycation in 3D Skin Models Leads to Alterations in Collagen Distribution and a Decrease in Stratification of the Epidermis
One of the biological effects of skin glycation is the accumulation of advanced glycation end products (AGEs) that can impair the collagen and elastic fibers in the dermis. To gain an insight into the effect of glycation on the cellular and molecular organization of the skin likely caused by such alterations, the 3D full-thickness skin models (1 cm in diameter, MatTek Life Sciences) were treated with reactive dicarbonyl compound Methylglyoxal (MGO) as described previously.26 MGO was included at a concentration of 500 mM in the growth media (a bottom chamber in contact with the dermis) for 7 days, and the accumulation of AGEs and pentosidine was initially measured by recording the auto-fluorescence of the models in a microplate reader at Ex/Em 370/440 nm and Ex/Em 335/385 nm on day 0, 1, 4 and 7. Auto-fluorescence as a non-invasive marker of AGEs accumulation in-vivo was described before.27,28 Treatment with MGO resulted in a rapid and significant increase in the signal at both Ex/Em wavelengths when normalized to the relative fluorescence intensity (RFI) from the untreated control. For Ex/Em 370/440 nm the RFI was increased by 2.2-fold, for Ex/Em 335/385 nm the RFI was increased by 1.3-fold; both signals reached the threshold between days 4 and 7 (Figure 1A). The increase in auto-fluorescence of the MGO-treated, unprocessed models could be additionally visualized under a fluorescence microscope with a filtration range at Ex/Em 350/460 nm corresponding to the blue (DAPI) filter (Figure 1B). These results indicated that MGO induced the formation of AGEs in skin models, which could be detected as auto-fluorescence and quantified.
 |
Figure 1 Glycation induced by MGO leads to the production of AGEs and alterations in collagen distribution in 3D skin models. (A) Skin models were cultured in the presence of methylglyoxal (MGO) and AGE-related auto-fluorescence measured in a microplate reader at Ex/Em 370/440 nm and Ex/Em 335/385 nm on days 0, 1, 4 and 7. Relative fluorescence intensity (RFI) in the control model equals 1, (B) Auto-fluorescence signals from the skin models under a fluorescence microscope, DAPI channel, (C) Photographs showing the diameter of the control and MGO-treated models, (D and E) Models were stained with the anti-CML antibody and H&E and the signal intensity of CML and eosin in the dermis quantified relative to control, (F) Masson’s trichrome staining of the models. Scale bars are 100 μm. p-values are *<0.05, **< 0.01, ***<0.001, n=3. |
Treatment with MGO also resulted in a noticeable decrease in the model diameter, which was reduced by ~20% at day 7 compared to untreated control, likely reflecting the alterations in mechanical properties due to modifications in the elastic fibers and extracellular matrix (ECM) in the dermis (Figure 1C). To gain further insight into the structural and cellular organization of the skin induced by glycation, the models were fixed, processed, and 5 mm thin sections prepared from wax-embedded samples. Staining with the antibody against N-carboxymethyl-lysine (CML), one of the major AGEs that causes glycoxidation and hallmarks damage to elastic fibers20,29 revealed a significant increase in the CML expression in the dermis of MGO-treated models compared to untreated control (Figure 1D). The sections were then stained with hematoxylin and eosin (H&E) and analyzed for the intensity of the eosin signal under the fluorescence microscope at Ex/Em 495/520 nm corresponding to the green (FITC) filter. Eosin fluorescence has been previously documented to reflect alterations in the cutaneous elastic fibers that can be used to evaluate the damage to the tissue.30 The analysis revealed a significant down-regulation in the fluorescent signal in the MGO-treated model, which was decreased by ~50% compared to the untreated control (Figure 1E). These data suggested a likely extensive re-organization and decrease in the density of collagen and elastic fibers due to MGO-induced glycation events in the dermis. To expand on this observation further, the sections were stained with Masson’s trichrome dye to obtain a more detailed structural overview of the tissue and to detect the density of the collagen fibers. In the control model, collagen was organized in dense bundles throughout the dermis; this was also accompanied by the presence of a defined collagen-dense cellular layer located directly underneath the epidermis. In the MGO-treated model, collagen was distributed more uniformly and visibly reduced in density in the upper-most dermis engaged in direct contact with the epidermis (Figure 1F, arrow).
These data indicated that glycation induces changes to the skin’s architecture that could also affect the specific compartments of the dermis involved in the signaling interactions with the epidermis. Dermal–epidermal interactions are crucial for the maintenance of inductive signals involved in skin homeostasis, including self-renewal and differentiation of keratinocytes leading to the formation of the stratified barrier.31 Quantification of the densities of nucleated cells in the dermal-epidermal area of skin sections stained with nuclear intercalating dye DAPI revealed a ~50% decrease in the densities of fibroblasts and ~50% increase in the densities of keratinocytes in the MGO-treated model compared to control (Figure 2A). The increased cell numbers in the epidermis did not, however, correspond to increased stratification in the glycated model. Analysis of the epidermis in H&E-stained slides revealed a significant decrease in the epidermal thickness, which was reduced by ~30% in the MGO-treated model compared to control, accompanied by a visibly diminished thickness of the stratum corneum. Thinning of the epidermis was associated with a ~60% decrease in the quantities of granular cells with flattened nuclear morphology, which normally contribute to the upper cornified layers through terminal differentiation (Figure 2B). Subsequent staining with the antibody against proliferation marker Ki67, detecting the dividing population in the basal cells layer, revealed a ~50% reduction in Ki67-positive cells in the MGO-treated model compared to control (Figure 2C and D). These results indicated that glycation leads to likely alterations in the epidermal structure and homeostasis, involving accumulation of the nucleated, slowly dividing keratinocytes with diminished stratification potential. Such events are also accompanied by changes in the cellular and molecular organization of the dermis.
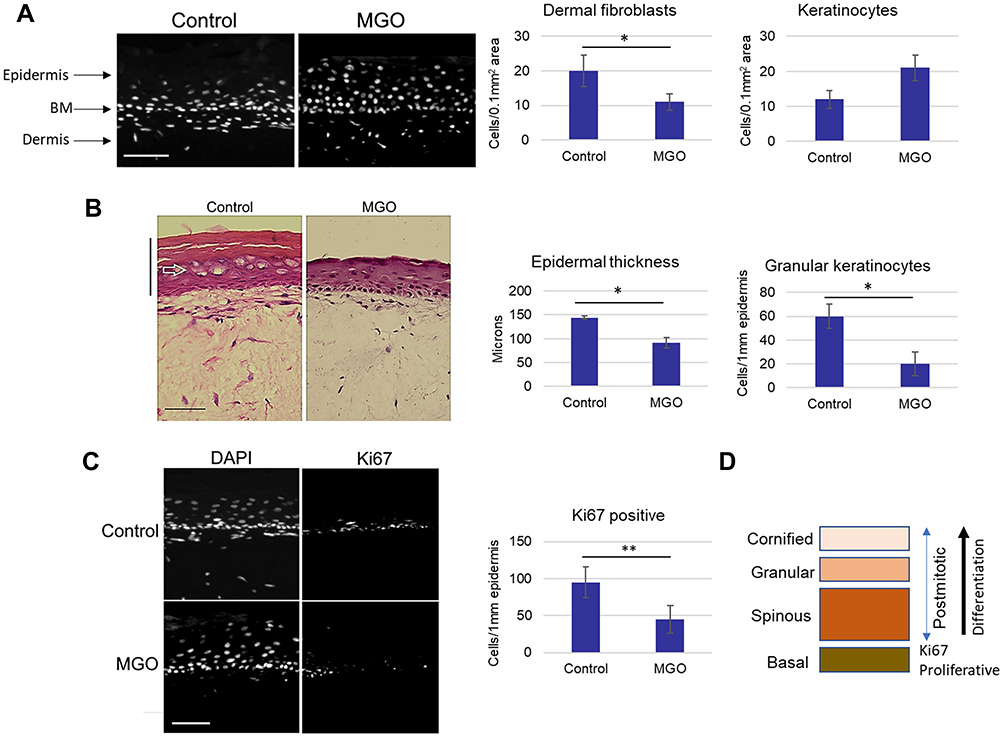 |
Figure 2 Glycation induced by MGO affects the kinetics of epidermalization. (A) Skin models were cultured in the presence of MGO and cell densities were quantified in the area of the upper dermis and the epidermis. (B) Epidermal thickness and the relative numbers of granular keratinocytes quantified from H&E- stained images of control and MGO-treated models, (C) Percentage of the Ki67-positive cells in the epidermis were calculated from the fluorescence images of control and MGO-treated models, (D) Schematic representation of the proliferative versus differentiating cellular populations in the epidermis. Scale bars are 100 μm. p-values are *<0.05, **< 0.01, n=3. |
Treatment of Dermal Fibroblasts with Resveratrol and Its Derivatives Reduces the Senescent Phenotypes Induced by Glycation
It is well established that alterations in the kinetics of epidermalization and organization of the elastic fibers are some of the characteristics of skin aging. Based on the observation that the treatment of skin models with MGO also appeared to affect the fibroblast density in the dermis, we sought to investigate the likely aging effect of glycation on dermal fibroblasts and the application of bioactive compounds with the potential capacity to ameliorate such impact on the cells. To address this, human dermal fibroblasts (HDFs) were cultured at young passage (p4-p5) and treated for 72 hours with 500 mM MGO in growth media. Fluorescence microscopy images captured at Ex/Em 350/460 nm revealed that MGO treatment increased the auto-fluorescence of the cells, which was absent in untreated control cells (Figure 3A). The auto-fluorescence signals could also be quantified in the microplate reader; at both Ex/Em 370/440 nm and Ex/Em 335/385 nm the relative fluorescence intensity was increased between ~75% and ~50% in MGO treated cells compared to control (Figure 3B).
 |
Figure 3 Senescent phenotypes of dermal fibroblasts cultured in the presence of MGO. (A) HDF cultures were treated with MGO for 72 hours and the auto-fluorescence signals were captured under a fluorescence microscope, DAPI channel, alongside phase contrast images; (B) AGE-related auto-fluorescence measured in a microplate reader at Ex/Em 370/440 nm and Ex/Em 335/385 nm. RFI in the control model equals 1; (C) The cell shapes and cell densities in control and MGO-treated cultures stained with crystal violet; (D and E) Percentage of the Ki67- and p16INK4a- positive cells calculated from the fluorescence images of control and MGO-treated cultures; (F) Intracellular ROS production measured using DCF-DA assay, relative DCF fluorescence at Ex/Em 485/535 nm in control equals 1. Scale bars are 50 μm. p-values are *<0.05, **< 0.01, n=3. |
The presence of MGO in culture media also induced significant alterations in cell morphology, specifically enlarged size and flattened shapes reminiscent of senescent appearance. These changes were accompanied by a decrease in cell densities, which were reduced by ~60% in MGO-treated HDF cultures compared to control (Figure 3C). Inhibition of cell divisions in the presence of MGO could be additionally confirmed by the quantification of the Ki67-positive cells; which were reduced by ~30% compared to control cells (Figure 3D). To correlate the effect of MGO on cell morphology and proliferative capacities with aging-related characteristics, we stained the HDF cultures with p16 INK4a, a cyclin-dependent kinase inhibitor, and the marker of cell senescence. Expression of p16 INK4a did show an increase by ~25% in the MGO-treated cultures compared to control (Figure 3E). Consistently, culturing the cells in the presence of MGO also resulted in a significant increase in the oxidative stress and upregulated intracellular production of ROS, which was increased by ~75% in the dichlorodihydrofluorescein diacetate (DCF-DA) assay (Figure 3F). These results indicated that MGO-induced glycation in dermal fibroblasts is also associated with a phenotype consistent with cellular aging, which includes alterations in cell morphology, cell cycle and an increase in oxidative stress.
To address whether such responses could be ameliorated to protect the skin cells from glycation-induced damage, we analyzed the anti-glycation potential and cellular effects of four bioactive compounds, Resveratrol (RESV) and its derivatives: OxyResveratrol (OxyRESV), Piceatannol (PICTL) and Triacetyl Resveratrol (TraRESV) in the MGO-treated fibroblast cultures. In addition, we also included Carnosine (CARN), a well-characterized compound with anti-glycation potential, in the analysis.20
Treatment of the fibroblasts with MGO and the compounds (0.01% concentration) for 72 hours revealed that all molecules were able to reduce the auto-fluorescence of the cultures measured in the microplate reader. For both Ex/Em 370/440 nm and Ex/Em 335/385nm, detecting the AGEs and the pentosidine, respectively, the reduction in auto-fluorescence was most pronounced for Resveratrol and OxyResveratrol and estimated at ~30% of the RFI from the MGO-treated control (Figure 4A).
 |
Figure 4 Treatment of dermal fibroblasts with Resveratrol and its derivatives reduces the senescent phenotypes induced by glycation. HDF cultures were treated with MGO or with MGO plus bioactive compounds: Carnosine, Resveratrol, OxyResveratrol, Piceatannol and TracetylResveratrol. (A) AGE-related auto-fluorescence measured in a microplate reader at Ex/Em 370/440 nm and Ex/Em 335/385 nm on day 7, (B) The cell shapes and cell densities in MGO- and compounds-treated cultures stained with crystal violet; Scale bars are 50μm, (C) Percentage of the p16INK4a- positive cells calculated from the fluorescence images of the cultures. (D) Intracellular ROS production in MGO- and compounds-treated cultures measured using DCF-DA assay, relative DCF fluorescence at Ex/Em 485/535 nm in untreated control equals 1, (E) The skin models were treated with MGO and topically applied compounds and AGE-related auto-fluorescence measured in a microplate reader at Ex/Em 370/440 nm and Ex/Em 335/385 nm at day 7, (F) The epidermal morphology in skin models sections stained with H&E. Scale bars are 100 μm. p-values are *<0.05, **< 0.01, n=3. |
All compounds also had the ability to prevent the appearance of enlarged, flattened cell morphology induced by glycation; the cells adopted more elongated shapes characteristic of the proliferative cultures. Preservation of cell shapes was also accompanied by a significant increase in cell densities, which was most pronounced for Resveratrol and OxyResveratrol (~6-fold compared to MGO-treated control, Figure 4B).
Staining of the cultures with the antibody against senescent marker p16 INK4a revealed a significant reduction in the expression levels for all compounds, which was reduced by ~75% compared to the MGO-treated control (Figure 4C). Consistent with the anti-aging effect, the compounds were also strongly inhibiting the intracellular ROS production in the DCF-DA assay, which was most pronounced for Carnosine, Resveratrol and OxyResveratrol (~50% DCF signal reduction compared to MGO-treated control, Figure 4D). These data suggested that Carnosine, Resveratrol and Resveratrol’s derivatives can reduce the effects of glycation on skin dermal fibroblasts; with several parameters including generation of AGEs, changes in proliferative capacities, expression of senescence marker and oxidative stress, indicative of the protective and anti-aging effect on the cells.
Dermal fibroblasts constitute an integral component of the skin, contributing not only to the structural integrity and elasticity of the dermis but also to signaling interactions responsible for epidermal homeostasis and renewal.31 To evaluate the anti-glycation activity and the effect on the epidermis, the compounds were subsequently tested in 3D skin models. The models were treated with the 500 mM MGO in growth media (bottom chamber) to induce glycation, simultaneously 0.5% aqueous solutions of Carnosine, Resveratrol, OxyResveratrol, Piceatannol and Triacetyl Resveratrol were applied topically, and the models were processed for analysis after 7 days. Consistent with the data from fibroblast cultures, all compounds also had the capacity to reduce the auto-fluorescence of skin models induced by MGO glycation. For both Ex/Em 370/440 nm and Ex/Em 335/385 nm measurements, the RFI was reduced by ~50% and was significant except for Triacetyl Resveratrol (4E). To correlate this activity with the possible effect on the epidermis, the models were processed and the H&E-stained sections were analyzed for a degree of stratification. The screening revealed that the epidermal thickness and stratification in the MGO-treated model were both enhanced in the presence of Carnosine, Resveratrol and OxyResveratrol, in contrast, the epidermalization was less pronounced when Piceatannol and Triacetyl Resveratrol were applied (Figure 4F).
These results suggested that Carnosine, Resveratrol and OxyResveratrol could not only inhibit the formation of AGEs but also had the most optimal epidermalization capacities in 3D skin models. In addition, these abilities were also strongly correlated with the anti-aging effect on dermal fibroblasts, with parameters such as preservation of cell densities most pronounced for Resveratrol and OxyResveratrol.
Topical Applications of Carnosine, Resveratrol and OxyResveratrol in Skin Models Ameliorate Structural Defects in the Dermis and Kinetics of Epidermalization Induced by Glycation
Having identified Resveratrol, OxyResveratrol and Carnosine as the most optimal compounds, we selected them for further analysis. Skin models were cultured in the presence of 500 mM MGO in growth media (bottom chamber) and treated simultaneously with 0.5% aqueous solutions of the compounds applied topically. The models were then analyzed for the auto-fluorescence at the intervals of 0, 1, 4, and 7 days. Both Resveratrol and OxyResveratrol were more effective in inhibition of AGEs accumulation induced by MGO than Carnosine, with the lower RFI values evident between day 4 and 7 for both Ex/Em 370/440 nm and Ex/Em 335/385 nm measurements. At Ex/Em 370/440 nm, the RFI was reduced by ~30% for Carnosine, ~50% for Resveratrol and ~60% for OxyResveratrol compared to the MGO-treated control. At Ex/Em 335/385 nm the OxyResveratrol also demonstrated a significant early reduction in RFI, by ~75% on day 1 indicative of a strong protective effect against pentosidine formation (Figure 5A). The decrease in the auto-fluorescence of the MGO-treated models in the presence of the compounds can also be visualized under the fluorescent microscope and filtration range at Ex/Em 350/460 nm corresponding to the blue (DAPI) filter (Figure 5B).
 |
Figure 5 Topical applications of Carnosine, Resveratrol and OxyResveratrol in skin models inhibit the formation of AGEs and protect against structural defects induced by glycation. (A) Skin models were treated with MGO and topically applied Carnosine, Resveratrol and OxyResveratrol; AGE-related auto-fluorescence was measured in a microplate reader at Ex/Em 370/440 nm and Ex/Em 335/385 nm at day 0, 1, 4 and 7. RFI in the control model equals 1, (B) Auto-fluorescence signals from the skin models under a fluorescence microscope, DAPI channel, (C) Models were stained with the anti-CML antibody and the signal intensity of CML in the dermis and the epidermis quantified relative to the MGO-treated control. Scale bars are 25 μm, (D) Models were stained with H&E and the signal intensity of eosin in the dermis was quantified relative to the MGO-treated control, (E) Masson’s trichome staining of the models. Scale bars are 50 μm. p-values are *<0.05, **< 0.01, ***<0.001, n=3. |
Further analysis of the model sections stained with the antibody against CML demonstrated that all compounds could also significantly inhibit its formation induced by MGO. Consistent with the auto-fluorescence measurements for the AGEs and pentosidine, the reduction of CML expression was also more pronounced for Resveratrol and OxyResveratrol than Carnosine. In the MGO-treated dermis, CML expression was reduced by ~50% for Carnosine, ~75% for Resveratrol, and ~85% for OxyResveratrol upon topical application in the skin model. In the epidermis, CML expression was reduced by ~30% for Carnosine, ~40% for Resveratrol, and ~70% for OxyResveratrol compared to MGO-treated control. These data indicated that the compounds were also relatively more efficient in inhibition of CML formation in the dermis (Figure 5C).
To evaluate the anti-glycation activities of the compounds in the context of potential protection against MGO-induced damage to the elastic fibers and collagen, the sections were stained with H&E and visualized for the intensity of the eosin signal under a fluorescence microscope at Ex/Em 495/520 nm corresponding to FITC filter. The analysis revealed that while eosin fluorescence in the glycated model was enhanced in the presence of all compounds, the signal was particularly strong for Resveratrol and OxyResveratrol, likely reflecting increased stratification of the epidermis. Quantification of the fluorescence signal in the dermis indicated a significant increase by ~2-fold for Carnosine, ~4-fold for Resveratrol and ~8-fold for OxyResveratrol, indicative of the likely extensive protection against MGO-induced damage to the collagen and elastic fibers (Figure 5D).
The increased density of collagen could also be revealed through staining with Masson’s trichrome dye. Compared to the MGO-treated control, the compounds increased the formation of the collagen-dense layer localized directly below the epidermis, which was most pronounced for OxyResveratrol (Figure 5E).
To correlate these changes with the densities of fibroblasts in the models, we quantified the number of nucleated cells in the dermal-epidermal area of skin sections stained with DAPI. The analysis revealed a significant increase in fibroblast densities in the upper dermis for all compounds, ranging from ~1.5-fold for Carnosine, ~2-fold for Resveratrol, and ~2.5-fold for OxyResveratrol compared to the MGO-treated control model. These changes were accompanied by a ~25% reduction in the densities of nucleated keratinocytes for all compounds, which was most significant for Carnosine (Figure 6A).
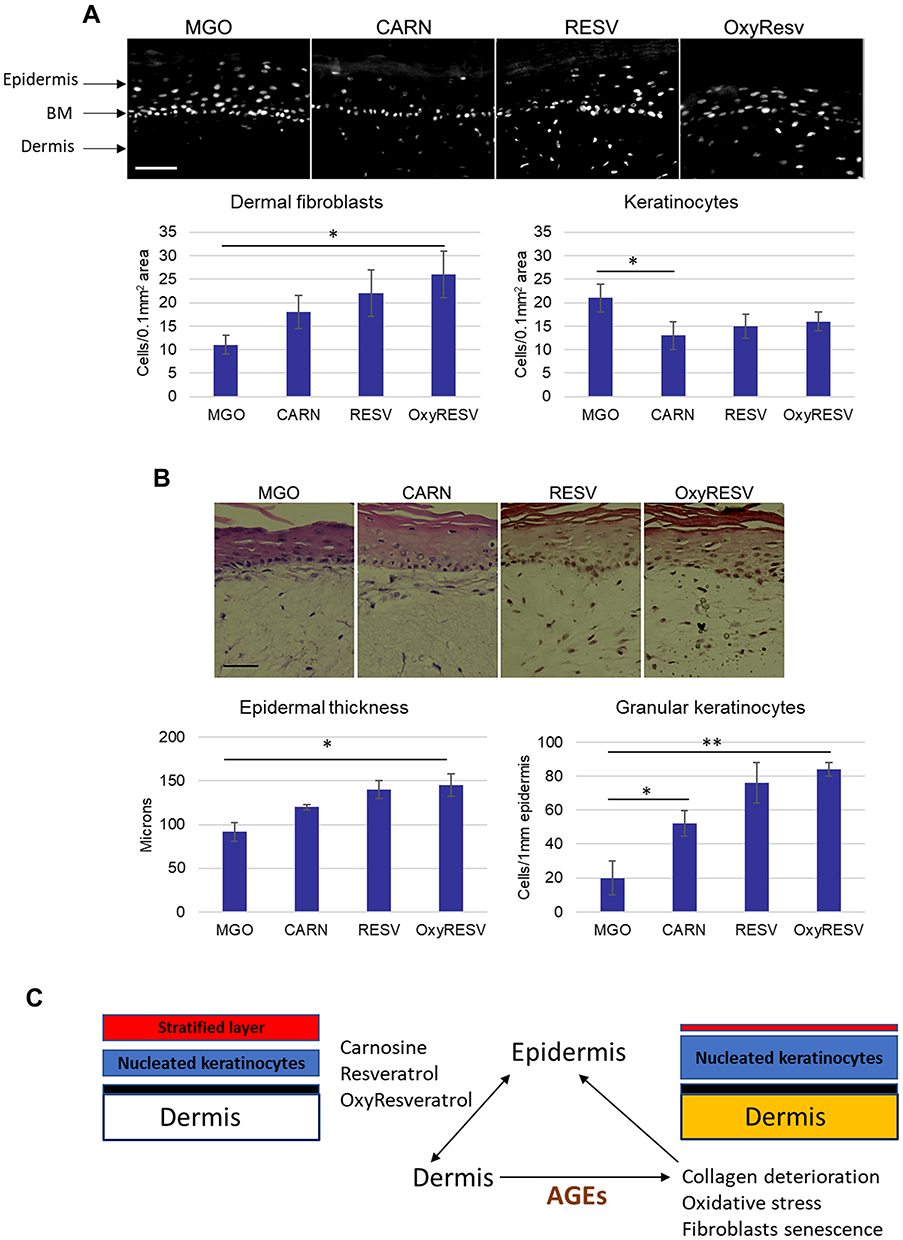 |
Figure 6 Topical applications of Carnosine, Resveratrol and OxyResveratrol in skin models improve the kinetics of epidermalization induced by glycation. (A) Skin models were treated with MGO and topically applied Carnosine, Resveratrol, and OxyResveratrol, and cell densities were quantified in the area of the upper dermis and the epidermis. Scale bars are 100 μm, (B) Epidermal thickness and the relative numbers of granular keratinocytes quantified from H&E- stained images of MGO-control and compounds-treated models. Scale bars are 50 μm; p-values are *<0.05, **< 0.01, n=3. (C) Schematic model summarizing the effect of MGO and the compounds on the dermis and epidermis in a 3D skin model. |
The decrease in the number of nucleated keratinocytes corresponded to increased stratification of the epidermis. The epidermal thickness in the MGO-treated model was increased by ~25% for Carnosine and ~50% for Resveratrol and OxyResveratrol, accompanied by the presence of cornified layers. These changes were associated with a significant increase in the quantities of granular cells with flattened nuclear morphology, indicative of a more efficient contribution to the upper cornified layers through terminal differentiation. Granular cells were increased by ~ 2.5 -fold for Carnosine and ~4-fold for Resveratrol and OxyResveratrol compared to the MGO-treated control, which was most pronounced in the presence of OxyResveratrol (Figure 6B).
These data indicated that Carnosine, Resveratrol and OxyResveratrol can prevent or restore the structural and cellular effects of MGO-induced glycation in skin models, namely the damage to the upper layers of dermis affecting collagen and fibroblast-dense region interacting directly with the epidermis; oxidative stress and alterations in the kinetics of epidermalization; evidenced by accumulation of nucleated keratinocytes with a diminished contribution to stratified layers forming skin barrier. These protective and anti-aging effects were moreover most optimal for Resveratrol and OxyResveratrol (Figure 6C, Summary model).
Discussion
This study has evaluated the effect of glycation on the skin based on the in-vitro 3D skin models; with a particular focus on the structural and cellular changes related to aging and the capacities of several natural compounds to prevent glycation-induced skin damage. We demonstrated that treatment of the models with glycation agent methylglyoxal (MGO) resulted in rapid accumulation of advanced glycation end products (AGEs), which could be detected as autofluorescence as described previously27,28 and quantified. The AGEs-associated fluorescence at Ex/Em 370/440 nm detects Amadori products involved in cross-linking interactions with larger molecules and formed early within the first few days post reaction.32 Pentosidine, measured in this study at Ex/Em 335/385 nm as described previously,27 is a glycoxidation product formed by sequential reactions of glycation and oxidation that progressively accumulate in skin and collagen, which is accelerated by high glycemic index.33 In our study, AGEs products measured at Ex/Em 370/440 nm could be detected early and increased steadily between days 1 and 4. In contrast, the quantities of pentosidine remained relatively low, most likely reflecting the longer-term requirement for physiological accumulation in the tissue.
Several polyphenols and flavonoids have been reported as promising anti-glycation candidates, including kaempferol, genistein, quercetin and epicatechin.22–24 The anti-glycation activity of known bioactive compounds can manifest through several mechanisms. First, they can interfere with the attachment of sugars to proteins and the formation of the Schiff base adduct in the initial stages of the glycation process. Secondly, they could prevent the formation of intermediate Amadori products and trapping of carbonyl compounds. Thirdly, they can inhibit the late stage of glycation by preventing the formation of AGEs from the intermediate products. Finally, a more indirect anti-glycation mechanism could also involve the ability to scavenge ROS in the antioxidant reactions. In the present study, we analyzed the anti-glycation activities of several bioactive compounds: Carnosine, Resveratrol and its derivatives, OxyResveratrol, Piceatannol and Triacetyl Resveratrol and found that the AGEs-related auto-fluorescence of skin models for both measurements at Ex/Em 370/440 nm and Ex/Em 335/385 nm was significantly reduced upon topical application of the compounds. Carnosine is a dipeptide molecule formed by beta-alanine and histidine and a natural compound produced by the body with strong anti-glycation properties documented previously.20 Resveratrol (3,4’,5-trihydroxystilbene) belongs to polyphenolic compounds stilbenes from plant sources, which has been proposed as a protective molecule with anti-glycation capacities.34 Both stilbenes, OxyResveratrol and Piceatannol have been demonstrated recently to exert a strong inhibitory effect on the formation of AGEs, and the anti-glycation capacity could be linked to the ability to scavenge methylglyoxal.35 Triacetyl Resveratrol, a synthetic and more stable derivative of Resveratrol, has the potential protective effects on the skin and can also affect the aging biomarkers.36,37 Topical application of the compounds in skin models demonstrated that the ability to inhibit AGEs formation induced by MGO, as measured by the auto-fluorescence, was most pronounced for Resveratrol, OxyResveratrol and Piceatannol. A subsequent time-course analysis could furthermore identify OxyResveratrol as the most potent for both reduction of AGEs at Ex/Em 370/440 nm and prevention of pentosidine formation at Ex/Em 335/385 nm. Consistent with these data, Resveratrol and OxyResveratrol also demonstrated the strongest inhibition of the formation of N-carboxymethyl-lysine (CML), a long-term glycoxidation product and one of the most common AGEs associated with damage to collagen and elastic fibers.29 This suggests that both compounds could have a potent protective effect on dermal architecture and skin elasticity through inhibition of AGEs formation that is linked to the glycemic index.
The analysis performed in this study uncovered likely extensive damage to the elastic fibers and collagen caused by MGO-induced glycation in skin models. Such damage was reflected by the decrease in the fluorescence signal of eosin in skin sections stained with hematoxylin and eosin (H&E), which could also bear some characteristics similar to the alterations in the cutaneous elastic fibers observed in photo-damage associated with premature aging.30 It is well established that in the skin, glycation causes cross-linking of AGEs in the extracellular matrix of the dermis, leading to loss of elasticity. Earlier studies on 3D skin models based on pre-glycated collagen have found increased activity of the ECM-degrading enzyme collagenase, suggesting enhanced collagen degradation that could be mechanistically linked to the changes accompanying aging.38 Such an observation is additionally supported by the identification of metalloproteinases and cytokines in pre-glycated collagen skin models, suggesting that the imbalance between ECM synthesis and degradation could drive glycation-induced skin aging.39 Our studies identified significant likely protection of Resveratrol and OxyResveratrol against the damage to the collagen and elastic fibers induced by MGO in the skin model.
Alterations in collagen density and organization caused by glycation were particularly extensive in the upper layer of the dermis, which remains in close proximal contact with the epidermis. A decrease in the fibrous collagen network in the upper dermis could be detected when the MGO-models were stained with Masson’s trichrome dye. These changes were also accompanied by a significant decrease in the density of dermal fibroblasts in the upper dermis compartment. Resveratrol and to a higher extent OxyResveratrol could protect against the loss of collagen fibers in the upper dermis through glycation; similarly, the compounds could also preserve the densities of dermal fibroblasts, which remained at the levels similar to that observed in the untreated control. These results show that the compounds can have a direct effect on the molecular and cellular organization of the skin through inhibition of glycation. Although such capacities could be identified for all three compounds, Resveratrol and OxyResveratrol were more potent than Carnosine.
Skin is a highly stratified organ that in addition to the dermal compartment also contains the complex epidermal layers contributing to the outermost barrier rendering protection against harmful environmental factors. Dermal–epidermal interactions are vital for the maintenance and renewal of the epidermis; it is currently thought that direct signaling from the dermal compartment is involved in the control of the epidermal progenitor cells.40 Skin keratinocytes are organized in well-defined populations, which migrate upwards and undergo sequential programs commencing with proliferation in the deepest basal layer, through cell cycle arrest and progressive differentiation to spinous and then to granular cells that eventually become enucleated and terminally differentiated to form stratum corneum in the uppermost part of the epidermis.31 Our study found that treatment of skin models with MGO also led to the accumulation of nucleated keratinocytes and decreased quantities of granular cells, which was associated with reduced thickness and stratification of the epidermis compared to control. These characteristics could be prevented by topical applications of Resveratrol and OxyResveratrol, which were again more potent than Carnosine, indicative of the strong protective capacities towards skin barrier formation and maintenance.
Loss of elasticity in the dermis and epidermal thinning are the important characteristics of skin aging that contribute to deteriorating quality in its appearance.41 It is presently unclear whether skin aging is caused by a sequence of events initiated in one specific compartment. The epidermis in aged skin has weakened barrier function caused by decreased turnover rate, changes in terminal differentiation and organization of the stratum corneum.42 It has been proposed that the deficiencies in differentiation capacity can stem from more abundant transit-amplifying (TA) cells with much longer cell cycle transit times.43 Increased quantities of nucleated keratinocytes accompanied by impaired stratification and decreased expression of the proliferation marker Ki67 observed in basal cells in the present study are consistent with aging phenotypes induced by skin glycation; these changes could be prevented by topically applied natural compounds including Carnosine, Resveratrol and OxyResveratrol.
Loss of skin elasticity and changes in the superficial dermis in proximity to the epidermis are the hallmarks of skin aging. This can be caused not only by decreased synthesis of collagen by dermal fibroblasts but also by fragmentation of the collagen matrix by metalloproteinases (MMPs), leading to a self-sustaining feedback loop and increase in fibroblast senescence.44,45 Aged skin is also characterized by increased production of ROS and reduced antioxidant activity.46 It is therefore likely that the accumulation of senescent fibroblasts in the dermis contributes to skin aging. Treatment of dermal fibroblasts with MGO performed in this study resulted in auto-fluorescence that could be captured under a fluorescence microscope and quantified in a microplate reader, reflective of the accumulation of AGEs relative to untreated control. The MGO-treated fibroblasts were also characterized by enlarged, flattened cell shapes and decreased cell densities, decreased expression of proliferation marker Ki67, increased expression of senescence marker p16INK4a and enhanced intracellular ROS production, consistent with the senescent phenotypes. Both ROS production and p16INK4a expression induced by MGO were significantly inhibited when the fibroblasts were cultured in the presence of Carnosine, Resveratrol, OxyResveratol, Piceatannol and Triacetyl Resveratrol. This effect could be associated with the general capacities of the compounds, which are well documented as anti-oxidants.47,48 Treatment with the compounds also prevented the alterations in cell shapes and reduction in cell densities induced by MGO, indicative of the potential capacities to inhibit the senescent phenotypes. Although cell densities in MGO-treated cultures were increased in the presence of all compounds, the effect was most pronounced for Resveratrol and OxyResveratrol and coincided with the most significant reduction in the Ex/Em 370/440 nm and Ex/Em 335/385 nm auto-fluorescence. These data suggest that the anti-aging potential of Resveratrol and OxyResveratrol could be linked to both robust anti-glycation activity and preservation of high fibroblast numbers in the skin dermis.
Cellular senescence has been shown to utilize the senescence-associated secretory pathway (SASP);49 it can be proposed that the cellular aging induced by glycation in the dermal environment also drives the epidermal thinning and loss of barrier function through the bystander effect. Natural compounds have an intrinsic poly-pharmacology effect that is attributed to the multiple biological activities linked to the likely capacities to modulate the bystander effect.50 An increase in fibroblasts densities in the upper dermis could be critical to supporting the signaling interactions with the epidermis; these specific anti-aging characteristics were most pronounced for Resveratrol and OxyResveratrol and could explain the most optimal capacity to protect the epidermis and barrier formation against glycation and damage.
Conclusion and Proposed Model
Skin glycation causes damage to the upper layers of the dermis, particularly collagen and fibroblast-dense region interacting directly with the epidermis, leading to changes reminiscent of senescence. Such events are closely associated with alteration in the kinetics of epidermalization; evidenced by the accumulation of nucleated keratinocytes with diminished differentiation capacity and contribution to stratified layers forming skin barrier. These effects of glycation can be prevented or restored by the natural compounds, with Resveratrol and more so OxyResveratrol demonstrating the strongest effect on inhibition of AGEs formation, increase in fibroblast density, inhibition of ROS production, collagen restoration and epidermal stratification. Topical applications of OxyResveratrol could have a novel and potent effect on anti-glycation and skin rejuvenation in cosmetic products.
Abbreviations
AGE, advanced glycation end products; CARN, Carnosine; CML, N-carboxymethyl-lysine; DAPI, 4ʹ6-diamidino-2-phenylindole; DCF-DA, dichlorodihydrofluorescein diacetate; DMEM, Dulbecco’s Modified Eagle’s Medium; ECM, extracellular matrix; EDTA, Ethylenediamine tetraacetic acid; FBS, fetal bovine serum; FITC, Fluorescein Isothiocyanate; HDF, human dermal fibroblasts; H&E, hematoxylin and eosin; MGO, Methylglyoxal; NOS, nitrogen species; OxyRESV, OxyResveratrol; PBS, phosphate-buffered saline; PICTL, Piceatannol; RESV, Resveratrol; RFI, relative fluorescence intensity; ROS, reactive oxygen species; TCA, tricarboxylic acid; TraRESV, Triacetyl Resveratrol; TRITC, Tetramethyl Rhodamine Iso-Thiocyanate.
Disclosure
Jaimie Jerome and Thomas Mammone are employees of Estee Lauder Inc. Ewa Markiewicz and Olusola C Idowu are employees of HexisLab Limited. The authors report no other conflicts of interest in this work.
References
1. Stopper H, Schinzel R, Sebekova K, Heidland A. Genotoxicity of advanced glycation end products in mammalian cells. Cancer Lett. 2003;190:151–156. doi:10.1016/s0304-3835(02)00626-2
2. Ahmed N, Thornalley PJ. Quantitative screening of protein biomarkers of early glycation, advanced glycation, oxidation and nitrosation in cellular and extracellular proteins by tandem mass spectrometry multiple reaction monitoring. Biochem Soc Trans. 2003;31:1417–1422. doi:10.1042/bst0311417
3. Boule NG, Haddad E, Kenny GP, et al. Effects of exercise on glycemic control and body mass in type 2 diabetes mellitus: a meta-analysis of controlled clinical trials. JAMA. 2001;286:1218–1227. doi:10.1001/jama.286.10.1218
4. Huebschmann AG, Regensteiner JG, Vlassara H, Reusch JE. Diabetes and advanced glycoxidation end products. Diabetes Care. 2006;29:1420–1432. doi:10.2337/dc05-2096
5. Sugiyama S, Miyata T, Ueda Y, et al. Plasma levels of pentosidine in diabetic patients: an advanced glycation end product. J Am Soc Nephrol. 1998;9:1681–1688. doi:10.1681/ASN.V991681
6. Semba RD, Nicklett EJ, Ferrucci L. Does accumulation of advanced glycation end products contribute to the aging phenotype? J Gerontol A Biol Sci Med Sci. 2010;65:963–975. doi:10.1093/gerona/glq074
7. Schlotterer A, Kukudov G, Bozorgmehr F, et al. C. elegans as model for the study of high glucose-mediated life span reduction. Diabetes. 2009;58:2450–2456. doi:10.2337/db09-0567
8. Wright KJ, Thomas MM, Betik AC, et al. Exercise training initiated in late middle age attenuates cardiac fibrosis and advanced glycation end-product accumulation in senescent rats. Exp Gerontol. 2014;50:9–18. doi:10.1016/j.exger.2013.11.006
9. van Deemter M, Ponsioen TL, Bank RA, et al. Pentosidine accumulates in the aging vitreous body: a gender effect. Exp Eye Res. 2009;88:1043–1050. doi:10.1016/j.exer.2009.01.004
10. Hu S, He W, Liu Z, et al. The accumulation of the glycoxidation product N(epsilon)-carboxymethyllysine in cardiac tissues with age, diabetes mellitus and coronary heart disease. Tohoku J Exp Med. 2013;230:25–32. doi:10.1620/tjem.230.25
11. Bakala H, Ladouce R, Baraibar MA, Friguet B. Differential expression and glycative damage affect specific mitochondrial proteins with aging in rat liver. Biochim Biophys Acta. 2013;1832:2057–2067. doi:10.1016/j.bbadis.2013.07.015
12. Roca F, Grossin N, Chassagne P, et al. Glycation: the angiogenic paradox in aging and age-related disorders and diseases. Ageing Res Rev. 2014;15:146–160. doi:10.1016/j.arr.2014.03.009
13. Ott C, Jacobs K, Haucke E, et al. Role of advanced glycation end products in cellular signaling. Redox Biol. 2014;2:411–429. doi:10.1016/j.redox.2013.12.016
14. Abdo JM, Sopko NA, Milner SM. The applied anatomy of human skin: a model for regeneration. Wound Med. 2020;28:100179. doi:10.1016/j.wndm.2020.100179
15. Nguyen HP, Katta R. Sugar sag: glycation and the role of diet in aging skin. Skin Ther Lett. 2015;20:1–5.
16. Pageon H, Bakala H, Monnier VM, Asselineau D. Collagen glycation triggers the formation of aged skin in vitro. Eur J Dermatol. 2007;17:12–20. doi:10.1684/ejd.2007.0102
17. Verzijl N, DeGroot J, Oldehinkel E, et al. Age-related accumulation of Maillard reaction products in human articular cartilage collagen. Biochem J. 2000;350(Pt 2):381–387. doi:10.1042/bj3500381
18. Avery NC, Bailey AJ. The effects of the Maillard reaction on the physical properties and cell interactions of collagen. Pathol Biol. 2006;54:387–395. doi:10.1016/j.patbio.2006.07.005
19. Nowotny K, Grune T. Degradation of oxidized and glycoxidized collagen: role of collagen cross-linking. Arch Biochem Biophys. 2014;542:56–64. doi:10.1016/j.abb.2013.12.007
20. Nardaa M, Peno-Mazzarinob L, Krutmann J, et al. Novel facial cream containing carnosine inhibits formation of advanced glycation end-products in human skin. Skin Pharmacol Physiol. 2018;31:324–331. doi:10.1159/000492276
21. Peng X, Ma J, Chen F, Wang M. Naturally occurring inhibitors against the formation of advanced glycation end-products. Food Funct. 2011;2:289–301. doi:10.1039/c1fo10034c
22. Kim J, Kim CS, Moon MK, Kim JS. Epicatechin breaks preformed glycated serum albumin and reverses the retinal accumulation of advanced glycation end products. Eur J Pharmacol. 2015;748:108–114. doi:10.1016/j.ejphar.2014.12.010
23. Lv L, Shao X, Chen H, et al. Genistein inhibits advanced glycation end product formation by trapping methylglyoxal. Chem Res Toxicol. 2011;24:579–586. doi:10.1021/tx100457h
24. Xie Y, Chen X. Structures required of polyphenols for inhibiting advanced glycation end products formation. Curr Drug Metab. 2013;14:414–431. doi:10.2174/1389200211314040005
25. Brownlee M, Vlassara H, Kooney A, et al. Aminoguanidine prevents diabetes-induced arterial wall protein cross-linking. Science. 1986;232:1629–1632. doi:10.1126/science.3487117
26. Sadowska-Bartosz I, Galiniak S, Bartosz G. Kinetics of glycoxidation of bovine serum albumin by methylglyoxal and glyoxal and its prevention by various compounds. Molecules. 2014;19:4880–4896. doi:10.3390/molecules19044880
27. Gugliucci A, Menini T. Circulating advanced glycation peptides in streptozotocin-induced diabetic rats: evidence for preferential modification of IgG light chains. Life Sci. 1998;62(23):2141–2150. doi:10.1016/s0024-3205(98)00189-1
28. Da Moura Semedo C, Webb M, Waller H, et al. Skin autofluorescence, a non-invasive marker of advanced glycation end products: clinical relevance and limitations. Post Med J. 2017;93:289–294. doi:10.1136/postgradmedj-2016-134579
29. Yoshinaga E, Kawada A, Ono K, et al. N(ɛ)-(carboxymethyl)lysine modification of elastin alters its biological properties: implications for the accumulation of abnormal elastic fibers in actinic elastosis. J Invest Dermatol. 2012;132(2):315–323. doi:10.1038/jid.2011.298
30. Heo YS, Song HJ. Characterizing cutaneous elastic fibers by eosin fluorescence detected by fluorescence microscopy. Ann Dermatol. 2011;23(1):44–52. doi:10.5021/ad.2011.23.1.44
31. Rognoni E, Watt FM. Skin cell heterogeneity in development, wound healing and cancer. Trends Cell Biol. 2018;28(9):709–722. doi:10.1016/j.tcb.2018.05.002
32. Wu JT, Tu MC, Zhung P. Advanced glycation end product (AGE): characterization of the products from the reaction between D-glucose and serum albumin. J Clin Lab Anal. 1996;10(1):21–34. doi:10.1002/(SICI)1098-2825(1996)10:1<21::AID-JCLA5>3.0.CO;2-O
33. Booth AA, Khalifah RG, Todd P, Hudson BG. In vitro kinetic studies of formation of antigenic advanced glycation end products (AGEs). Novel inhibition of post-Amadori glycation pathways. J Biol Chem. 1997;272:5430–5437. doi:10.1074/jbc.272.9.5430
34. Maleki V, Foroumandi E, Hajiizadeh-Sharafabad F, et al. The effect of resveratrol on advanced glycation end products in diabetes mellitus: a systematic review. Arch Physiol Biochem. 2020;3:1–8. doi:10.1080/13813455.2019.1673434
35. Wang W, Yang R, Yao H, et al. Inhibiting the formation of advanced glycation end-products by three stilbenes and the identification of their adducts. Food Chem. 2019;295:10–15. doi:10.1016/j.foodchem.2019.02.137
36. Park J, Park JH, Suh HJ, et al. Effects of resveratrol, oxyresveratrol, and their acetylated derivatives on cellular melanogenesis. Arch Dermatol Res. 2014;306:475–487. doi:10.1007/s00403-014-1440-3
37. Hsieh TC, Wong C, Bennett DJ, Wu JM. Regulation of p53 and cell proliferation by resveratrol and its derivatives in breast cancer cells: an in silico and biochemical approach targeting integrin avb3. Int J Cancer. 2011;129:2732–2743. doi:10.1002/ijc.25930
38. Pageon H, Asselineau D. An in vitro approach to the chronological aging of skin by glycation of the collagen: the biological effect of glycation on the reconstructed skin model. Ann NY Acad Sci. 2005;1043:529–532. doi:10.1196/annals.1333.060
39. Pageon H. Reaction of glycation and human skin: the effects on the skin and its components, reconstructed skin as a model. Pathol Biol. 2010;58(3):226–231. doi:10.1016/j.patbio.2009.09.009
40. Jahoda CA, Reynolds AJ. Hair follicle dermal sheath cells: unsung participants in wound healing. Lancet. 2001;358:1445–1448. doi:10.1016/S0140-6736(01)06532-1
41. Zouboulis CC, Makrantonaki E. Clinical aspects and molecular diagnostics of skin aging. Clin Dermatol. 2011;29:3–14. doi:10.1016/j.clindermatol.2010.07.001
42. McCullough JL, Kelly KM. Prevention and treatment of skin aging. Ann NY Acad Sci. 2006;1067:323–331. doi:10.1196/annals.1354.044
43. Charruyer A, Barland CO, Yue LL, et al. Transit-amplifying cell frequency and cell cycle kinetics are altered in aged epidermis. J Invest Dermatol. 2009;129:2574–2583. doi:10.1038/jid.2009.127
44. Fisher GJ, Quan T, Purohit T, et al. Collagen fragmentation promotes oxidative stress and elevates matrix metalloproteinase-1 in fibroblasts in aged human skin. Am J Pathol. 2009;174:101–114. doi:10.2353/ajpath.2009.080599
45. Varani J, Dame MK, Rittie L, et al. Decreased collagen production in chronologically aged skin: roles of age-dependent alteration in fibroblast function and defective mechanical stimulation. Am J Pathol. 2006;168:1861–1868. doi:10.2353/ajpath.2006.051302
46. Kohen R. Skin anti-oxidants: their role in aging and in oxidative stress- new approaches for their evaluation. Biomed Pharmacother. 1999;53:181–192. doi:10.1016/S0753-3322(99)80087-0
47. Prokopieva VD, Yarygina EG, Bokhan NA, Ivanova SA. Use of carnosine for oxidative stress reduction in different pathologies. Oxid Med Cell Longev. 2016;2016:2939087. doi:10.1155/2016/2939087
48. Leonard SS, Xia C, Jiang BH, et al. Resveratrol scavenges reactive oxygen species and effects radical-induced cellular responses. Biochem Biophys Res Commun. 2003;309(4):1017–1026. doi:10.1016/j.bbrc.2003.08.105
49. Davalos AR, Coppe JP, Campisi J, et al. Senescent cells as a source of inflammatory factors for tumor progression. Cancer Metastasis Rev. 2010;29:273–283. doi:10.1007/s10555-010-9220-9
50. Markiewicz E, Idowu OC. DNA damage in human skin and the capacities of natural compounds to modulate the bystander signalling. Open Biol. 2019;9(12):190208. doi:10.1098/rsob.190208
 © 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.