")
Back to Journals » Drug Design, Development and Therapy » Volume 16
The Beneficial Effects of Geniposide on Glucose and Lipid Metabolism: A Review
Received 14 July 2022
Accepted for publication 22 September 2022
Published 30 September 2022 Volume 2022:16 Pages 3365—3383
DOI https://doi.org/10.2147/DDDT.S378976
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Tuo Deng
Siting Gao,1– 3 Qin Feng1– 3
1Institute of Liver Diseases, Shuguang Hospital Affiliated to Shanghai University of Traditional Chinese Medicine, Hepatopathy Building, Shanghai, People’s Republic of China; 2Shanghai Key Laboratory of Traditional Chinese Clinical Medicine, Shuguang Hospital Affiliated to Shanghai University of Traditional Chinese Medicine, Shanghai, People’s Republic of China; 3Key Laboratory of Liver and Kidney Diseases, Ministry of Education, Shanghai University of Traditional Chinese Medicine, Shanghai, People’s Republic of China
Correspondence: Qin Feng, Institute of Liver Diseases, Shuguang Hospital Affiliated to Shanghai University of Traditional Chinese Medicine, 1200 Zhangheng Road, Pudong District, Shanghai, People’s Republic of China, Tel +86 13764380682, Fax +86 021-51322043, Email [email protected]
Abstract: Geniposide is a naturally sourced active ingredient that has diverse pharmacological effects and great potential in improving or treating different kinds of diseases. In recent years, more and more studies have confirmed that geniposide can improve glucose and lipid metabolism disorder, which is an increasingly prevalent health problem causing various metabolic diseases globally. Our review aims to summarize basic information on the pharmacological effects of geniposide on glucolipid metabolism. Geniposide increases glucose utilization and insulin production, protects pancreatic islet β cells, inhibits insulin resistance and hepatic glucose production, and suppresses gluconeogenesis. While in the aspect of lipid metabolism, geniposide can promote lipolysis, inhibit lipogenesis, and regulate lipid transport. Geniposide ameliorates lipid and glucose metabolic disorders, improving the entire glycolipid metabolism network in a three-dimensional manner at the level of molecular mechanism. Growing evidence revealed that geniposide may serve as an effective drug to combat metabolic diseases for the time to come.
Keywords: geniposide, lipid metabolism, glucose metabolism, Gardenia Jasminoides Ellis, metabolic disease, glucolipid metabolism, naturally sourced active ingredient, pharmacological evidence
Introduction
Glucose and lipid metabolism imbalance is a high-risk etiology leading to various complications, including obesity, diabetes, hyperlipemia, non-alcohol fatty liver disease (NAFLD), and cardiovascular diseases.1 We are currently in the midst of a global metabolic disease epidemic, with its prevalence increasing with age.2,3 There has been an increasing interest in investigating effective strategies to control and treat comorbidities associated with glucose and lipid metabolism disorders.4 A variety of plants and natural active ingredients derived from plants have been used to combat diseases associated with glycolipid metabolism disorders.5 Geniposide is one such naturally active ingredient derived from the fruits of Gardenia Jasminoides Ellis (GJE, popularly called Zhizi in China) and has traditionally been commonly used for hundreds of years in traditional Chinese medicine.6
There is an increasing number of pharmacological evidence proving that geniposide exerts various biological activities, including neuroprotective, antidiabetic, hepatoprotective, anti-inflammatory, analgesic, antidepressant-like, cardioprotective, antioxidant, immune-regulatory, antithrombotic, and anti-tumoral effects.7–11 The anti-inflammatory, hepatoprotective, antidiabetic, and antioxidant properties of geniposide were reviewed before.9,10,12–15 The most recognized pharmacological effects of geniposide are anti-inflammatory and antioxidant effects;9,13 however, in recent years, a growing number of studies explored the role of geniposide in regulating glucolipid metabolism. How geniposide regulates the glucolipid metabolism, the specific mechanism, the disadvantages and future research directions have not been elucidated. There are few reviews on the improvement of glucolipid metabolism disorders and metabolic diseases using geniposide. As a consequence, a comprehensive review focusing on geniposide-regulated glucolipid metabolism is necessary to advance the knowledge on geniposide. This review will serve as a reference for researchers to further understand the pharmacological effects of geniposide and develop more valuable applications in the future.
Geniposide at a Glance
Structure and Physicochemical Properties
Geniposide (methyl (1S,4aS,7aS)-1-(β-D-glucopyranosyloxy)-7-(hydroxylmethyl)-1,4a,5,7a-tetrahydrocyclopenta[c]pyran-4-carboxylate; C17H24O10) is regarded as an iridoid glycoside, namely genipin 1-O-β-D-glucopyranoside (Figure 1).10 From another perspective, geniposide is also regarded as a C11 methyl ester of geniposidic acid. Its molecular weight is 388,366. Because of its chemical structure, it is also considered a glycoside containing one molecule of genipin and glucose. Geniposide is a white or pale yellow powder with a density of 1.49 g/cm3 and a melting point of 161.53°C.7 It is easily dissolved in water, soluble in ethanol, and undissolved in petroleum ether.6
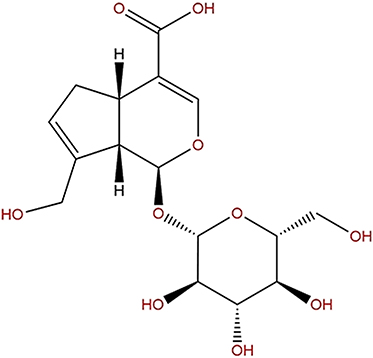 |
Figure 1 Chemical structure of geniposide. |
Sources
Geniposide is mainly sourced from GJE, but it is also detected in other commonly used Chinese herbal medicines, such as Eucommia Ulmoides Oliv (EUO), Rehmannia Officinalis (RO), and Radix Scrophulariae (RS) (Figure 2).16 Among these, GJE contains about 3.3–8.56% geniposide, while RO only contains 0.205–0.4381%, EUO contains 0.0173–0.5811%, and RS contains 0.0699–0.1135% of geniposide. Therefore, GJE is the main geniposide source.17 The 2015 edition of the Chinese Pharmacopoeia stipulates that geniposide abundance in GJE is no less than 1.8%. Geniposide content in GJE on the market reaches 6%.18
 |
Figure 2 Main sources of geniposide. (A). Gardenia Jasminoides Ellis (B). Eucommia Ulmoides Oliv (EUO) (C). Rehmannia Officinalis (RO) (D). Radix Scrophulariae (RS). |
Pharmacokinetics
We investigated that the concentration of geniposide reached a peak in the liver and spleen 30 min after oral administration, and was detected in the kidney and brain 2 h later.19 The absolute bioavailability of geniposide was 9.67% after it was metabolized within 12 h. The bioavailability of geniposide is strongly linked to different administration methods. In addition, geniposide can be converted to genipin by an intestinal microbiota enzyme (β-glucosidase) (Figure 3). Geniposide metabolism involves methylation, glucosylation, decarboxylation, taurine conjugation, hydrolysis, demethylation, hydrogenation, hydroxylation, cysteine S-conjugation through dehydration, sulfate conjugation, and other related complex reactions.20,21 Geniposide is mainly excreted in its original form through the kidneys. The excretion level accounts for a proportion of over 90% of the initial dosage after 10 h.22
 |
Figure 3 Schematic diagram of the metabolic effect of geniposide by β-glucosidase and esterase. |
Main Pharmacological Functions of Geniposide
Geniposide exerts abundant and complicated pharmacological effects, such as anti-inflammatory, antioxidant, hepatoprotective, neuroprotective, analgesic, antidiabetic, antidepressant-like, immune-regulatory, cardioprotective, antithrombotic, and antitumoral effects.8 These pharmacological effects have laid a foundation for its application in the improvement of a variety of diseases, such as cardiovascular diseases, diabetes and diabetic complications, hepatic diseases, Parkinson’s disease, Alzheimer’s disease, and ischemia and reperfusion injury. Some of the potential benefits include the influence on the normal and healthy operation of the nervous, endocrine, circulatory, digestive, urinary, muscle, and other different bodily systems.
In particular, the anti-inflammatory and antioxidant effects of geniposide have been widely studied. Geniposide not only fights inflammation-related diseases like swelling, pain, liver disease, Alzheimer’s disease, and others but also inhibits classic inflammatory-related pathways such as NF-κB, MAPK, and TLR4 signaling pathways.10 Geniposide could delay cell injury via upregulating endogenous antioxidative enzymes. Geniposide can also increase the activity of some important antioxidant enzymes and pathways including hepatic lipid peroxidation (LPO), glutathione-S-transferase (GST), glutathione (GSH), glutathione peroxidase (GPx), and copper- and zinc-containing superoxide dismutase (CuZn-SOD), and protects against oxidative stress injury.23–25
Pharmacological Effects of Geniposide in Glucose and Lipid Metabolism
Effects on Glucose Metabolism
It is well-known that an imbalanced glucose metabolism triggers many metabolic diseases. An adequate blood glucose level must be maintained at all time. Sources of blood glucose are intestinal absorption, liver glycogenolysis, and gluconeogenesis.26 Glucose-consuming pathways include glucose uptake by various tissues and organs for oxidation, glycogen synthesis, and conversion into other sugars, fats, or amino acids. The balance of blood glucose is mainly regulated by hormones, including insulin, glucagon, adrenaline, and glucocorticoid.27 The interaction between major glucose metabolic pathways, such as glycolysis, gluconeogenesis, glycogenesis, and glycogenolysis, maintains the homeostasis of hepatic glucose metabolism.28 Figure 4 summarizes the pharmacological effects of geniposide on glucose metabolism and the details are exhibited in Table 1.
 |  |  |  |
Table 1 In vivo/In vitro Studies of Geniposide in Glucose Metabolism |
 |
Figure 4 Pharmacological effects of geniposide on Glucose Metabolism. |
Increasing Glucose Utilization
Skeletal muscle, which is the main organ participating in the uptake and metabolism of glucose, contains slow-twitch and fast-twitch muscle fibers. Fast-twitch fibers generate adenosine triphosphate (ATP) primarily through glycolysis, whereas slow-twitch myofibers rich in mitochondria have high oxidative capacity.29 Geniposide improves glucose homeostasis by promoting a slow-to-fast myofiber switch and glucose utilization. Further studies exposed that geniposide exerts the above effects by regulating forkhead box O1 (FoxO1)/ pyruvate dehydrogenase kinase 4 (PDK4), which controls respiratory substrate selection through pyruvate dehydrogenase.30 From another point of view, in a 2022 study, it was revealed that geniposide regulates respiratory substrate selection, promotes glucose uptake in skeletal muscles, and suppresses glycogen storage by disturbing the synthesis, secretion, and homeostasis of retinol-binding protein 4 (RBP4).31
Increasing Insulin Production
Glucose-stimulated insulin secretion (GSIS) is essential to the maintenance of a stable level of blood glucose.32 Guo et al33 first reported that geniposide could increase GSIS, and the results indicated that glucagon-like peptide 1 receptors (GLP-1R) plays an important role in the geniposide-regulated GSIS. Geniposide regulates GSIS possibly via pyruvate carboxylase-mediated glucose metabolism in pancreatic β cells.34 Exploration from another point of view revealed that GSIS is phosphatidylinositol 3 kinase- dependent and geniposide increases the expression of glucose transporter 2 (GLUT2) in total cell lysates under normal glucose conditions.35 An in-depth study focusing on GLUT2, indicated that it may be related to the regulation of glucosaminyl (N-acetyl) transferase family member 7 (GNT-IVa)-mediated glycosylation of GLUT2 and the residence of glycosylated GLUT2 on the pancreatic β cell membrane.36 A 2021 study found that 5’AMP-activated protein kinase (AMPK) activation also plays an essential role in geniposide-regulated GSIS in pancreatic β cells.37
Protecting Pancreatic Islet β Cells
Diabetes is characterized by pancreatic islet β cell dysfunction or loss. One therapeutic strategy for improving blood glucose homeostasis is to prevent pancreatic islet β cells from failure and promote new pancreatic islet β cell formation.27
Geniposide is a promising pancreatic islet β cell protector, which prevents pancreatic islet β cells from exhaustion and injury resulting from excessive insulin secretion under high glucose conditions. Geniposide administration is a possible way of balancing the oxidative stress of pancreatic β cells by regulating the expression of protein disulfide isomerase (PDI) and endoplasmic reticulum oxidoreductin 1 (ERO1).38 Extensive islet amyloid polypeptide (IAPP) deposits are thought to contribute to pancreatic β cell dysfunction, either by direct cytotoxicity or by reducing the pancreatic islet β cell mass, resulting in impaired insulin secretion. Geniposide prevents human IAPP-induced cytotoxicity in INS-1E cells through the upregulation of (insulin degrading enzyme) IDE.39 Liu et al40 suggested that AMPK plays a crucial role in how geniposide antagonizes high glucose-induced pancreatic β cell injury.
Elevated thioredoxin-interacting protein (Txnip) levels induce β cell apoptosis and dysfunction.41 Geniposide improves GSIS by accelerating Txnip degradation. A further study proved that geniposide-related Txnip degradation attenuates the early-stage apoptosis of pancreatic β cells. Geniposide regulates Txnip degradation and GSIS through endoplasmic reticulum (ER) stress by accelerating the phosphorylation of Protein kinase R (PKR)-like endoplasmic reticulum kinase (PERK) and Inositol-requiring enzyme-1α (IRE1α).42
The main roles of GLP-1R are stimulation of GSIS, induction of pancreatic β-cell proliferation, inhibition of postprandial glucagon release, and delay in gastric emptying.43 Geniposide inhibits the early stage of lipotoxicity-induced β-cell apoptosis, and GLP-1R plays a critical role by counteracting lipotoxicity in INS-1 pancreatic islet β cells.44 Cui et al45 analyzed the microarray data of INS-1 cells treated with geniposide and identified key lncRNAs and mRNAs in a 2021 study. They found that the lncRNA NONRATT027738 interacts with all three hub mRNAs (Pomc, Htr2a, and Agtr1a). There is also other research focusing on the role of GLP-1R in the beneficial effects of geniposide-medicated pancreatic islet β cell protection. Geniposide potentiates insulin secretion via activating GLP-1R and adenylyl cyclase/cAMP signaling pathways. Researchers also observed Ca+2 channel activation by geniposide.39
Apart from inhibiting the damage and apoptosis of pancreatic islet β cells, another therapeutic strategy for the protection of pancreatic β cells is to promote their regeneration. Geniposide promotes pancreatic islet β cell regeneration in vivo to balance blood glucose levels and the mechanism includes triggering duct cell differentiation by enhancing TCF7L2 expression and activating the Janus-activated kinase 2 (JAK2) / signal transducer and activator of transcription 3 (STAT3) pathway.46
Improving Insulin Resistance (IR)
IR is an important factor, which can lead to the onset of type 2 diabetes. IR occurs when cells in muscles, fat, and liver stop responding to insulin. Geniposide alleviates abnormal glucose tolerance and hyperinsulinemia, indicating that it has an IR-alleviating effect.47 Geniposide promotes autophagy in HepG2 cells and significantly improves IR, which may be associated with the dynamic regulation of the P62/NF-κB/GLUT-4 pathway.48 Zhao et al49 pointed out that geniposide improves insulin signaling deficiency possibly through AMPK-mediated Txnip degradation in 3T3-L1 adipocytes. A regulatory mechanism study considered that geniposide could improve systemic insulin sensitivity by regulating circulating RBP4 levels.31
Inhibiting Hepatic Glucose Production
The liver plays a crucial role in maintaining blood glucose homeostasis by coordinating glucose storage, utilization, and production. The ability of the liver to store glycogen is typically diminished in diabetic subjects. Thus, promoting hepatic glycogen synthesis and suppression of hepatic glucose production could be more effective in the improvement of overall glycemic control. Geniposide significantly inhibits hepatic glucose production in a dose-dependent manner and the inhibitory effect is partly through AMPK activation.50 Geniposide simultaneously stimulates glycogen synthesis in mice induced by a high-fat diet and streptozotocin injection and HepG2 cells.
Suppressing Gluconeogenesis
Hepatic gluconeogenesis is an important factor in regulating plasma glucose levels. It may be the major source of fasting blood glucose and it has been regarded as one of the main contributors to hyperglycemia in diabetes mellitus. Phosphoenolpyruvate carboxykinase (PEPCK) and glucose-6-phosphatase (G6Pase) are the key regulatory enzymes of gluconeogenesis.50 Geniposide significantly decreases the expression of glycogen phosphorylase and G6Pase at mRNA and protein levels, as well as their activity, in a dose-dependent manner.51 The activities of PEPCK and G6Pase were significantly suppressed by geniposide. Geniposide may reduce blood glucose levels and suppress hepatic gluconeogenesis by regulating the AKT serine/threonine kinase (AKT)-FOXO1 pathway.52
Effects on Lipid Metabolism
Lipid metabolism disorder is one of the main risk factors of metabolic diseases, which is characterized by abnormal content or type of triglycerides, fatty acids, or cholesterol in the serum or surrounding tissues.53 Lipid metabolism includes three main aspects as follows: Lipolysis, lipogenesis, and lipid transportation. Regulating lipid metabolism and restoring lipid homeostasis is one of the methods for the prevention and treatment of dyslipidemia and related metabolic diseases.54 Figure 5 summarizes the pharmacological effects of geniposide on lipid metabolism and the details are exhibited in Table 2.
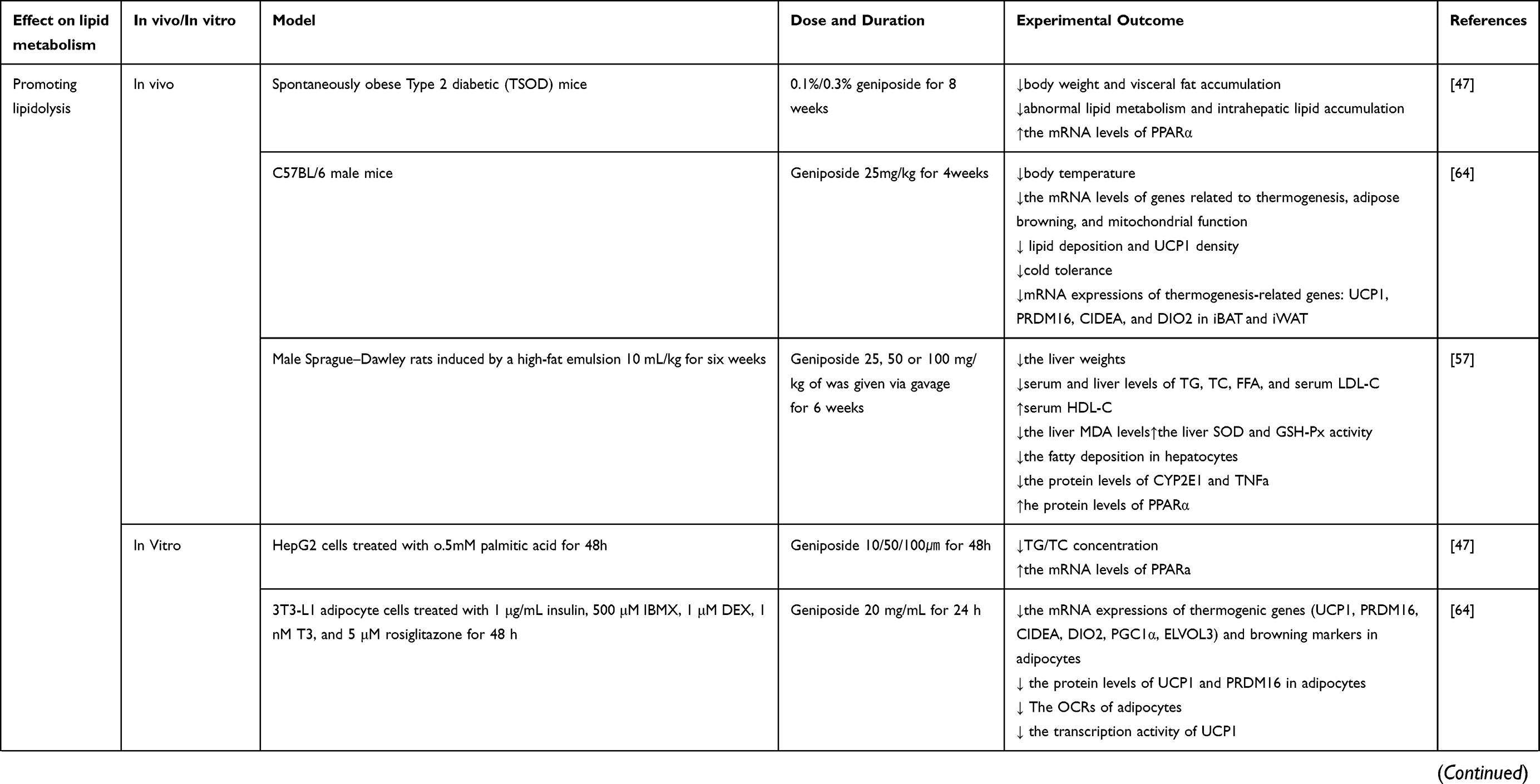 |  |  |
Table 2 In vivo/In vitro Studies of Geniposide in Lipid Metabolism |
 |
Figure 5 Pharmacological effects of geniposide on Lipid Metabolism. |
Promoting Lipolysis
Fat is mostly stored in adipose and other tissues or other non-adipose tissues. Excessive accumulation of triacylglycerols (TAGs) and cholesterol esters (CEs) leads to abnormal lipid metabolism. The lipid droplets (LDs) in the cytoplasm are mainly rich in TAGs and CEs, which are considered dynamic TAG storage pools and take part in several aspects of lipid metabolism.55
The primary oxidative pathway for energy production in the liver is β-oxidation which takes place in the mitochondria, the predominant regulators of which are the transcription factors peroxisome proliferators-activated receptors α (PPARα) and peroxisome proliferator-activated receptor gamma coactivator 1 alpha (Pgc1α).56 Geniposide exerts suppressive effects on hepatic lipid accumulation in rats fed with a high-fat diet and the underlying mechanism may be related to the regulation of adipocytokine release and PPARα expression.57 Geniposide improves the expression of NF-E2-related factor 2 (Nrf2), PPARγ, PPARα, and heme oxygenase 1 (HO-1) and regulates the AMPK/mTORC signaling pathway in mice. Genipin, the aglycone of geniposide, inhibits intracellular lipid accumulation and significantly increases PPARα mRNA expression.47 In the mitochondrion, FAs are oxidatively decomposed by carnitine palmitoyltransferase 1 (CPT1) to provide energy for other physiological processes. Sirtuin 1 (SIRT1), FoxO1, and PGC1α also control the activity of the related enzymes. Geniposide treatment alters the expression of FoxO1, PDK4, p-PDH and PDH at mRNA/protein level in the gastrocnemius of HFD-fed mice.31 In agreement with this finding, the inhibitory effects of geniposide on the FoxO1/PDK4/PDH signaling pathway have also been reported.30
Lipophagy (lysosome-mediated autophagy) and cytoplasmic lipolysis are two pathways to decompose TAGs and CEs in LDs. Geniposide increases the levels of autophagy in plaque macrophages via inhibiting the triggering receptors expressed on myeloid cells 2 (TREM2)/mechanistic target of rapamycin kinase (mTOR) axis.58 It may block the development of atherosclerosis through this mechanism. The more widely studied effect of geniposide on inflammation and oxidative stress through enhancing autophagy has been extensively studied recently, although further research is warranted.59–63
Activation of thermogenic adipocytes has great significance in the treatment of lipid metabolism disorders and is related to plenty of metabolic diseases. There is a new study published in 2021 different from the previous studies on the effect of geniposide on lipid metabolism. Thermogenic adipocyte is acknowledged to be a major regulator of energy homeostasis by affecting energy expenditure and glucolipid metabolism. Li et al64 suggested that geniposide is an inhibitor of fat thermogenesis in adipocytes through regulating the PKA signaling pathway, indicating that adipocyte thermogenic capacity may be inessential for geniposide to exert its effects in obesity, and the metabolic advantage of geniposide exerted on other organs, except for adipose tissues, can indemnify for the effect of geniposide-induced thermogenic activity of adipocytes. Further research on how geniposide regulates lipolysis through influencing thermogenesis in adipocytes is warranted.
Inhibiting Lipogenesis
Excessive lipid synthesis is a major cause of lipid metabolism disorders. The de novo lipogenesis, FAs obtained from acetyl-CoA to fatty acyl-CoA transformation, is principally regulated by acetyl-CoA carboxylase (ACaCa) and fatty acid synthase (FASN). The central enzymatic node of saturated fatty acids (SFAs) transformed into monounsaturated fatty acids (MUFAs) is stearoyl-CoA desaturase 1 (SCD1), which is regulated by sterol regulatory element-binding transcription factor 1 (SREBP-1c), carbohydrate-responsive element-binding protein (chREBP), and LXR. Transcriptional regulation of Acc and FASN happens mainly via SREBP-1c and chREBP. Geniposide is highly effective in inhibiting lipogenesis and free fatty acid (FFA) levels in hepatic tissues were decreased significantly after geniposide treatment, whereas TG, FASN, ACaCa, and malonyl-CoA levels were significantly reduced in the geniposide group.65 Geniposide could significantly suppress hepatic total triglyceride (TG), total cholesterol (TC), and FFA in NAFLD rodents.66 Geniposide also decreases the expression of SREBP-1c.67
Cholesterol plays a central role in lipid metabolism. TC and/or low-density lipoprotein cholesterol (LDL-c) levels ultimately depend on cholesterol hypersecretion. Significantly, geniposide inhibited TC and LDL production in different animal models. High mobility group box 1 (HMGCR), squalene monooxygenase, and SREBP are all crucial players in cholesterol synthesis (the first two are rate-limiting enzymes in the biosynthetic pathway, and the latter is a master transcriptional regulator of cholesterol synthesis). Geniposide inhibited TC, TG, LDL, and very-low-density lipoprotein (VLDL) production, SREBP1-c expression, and HMGCR expression at the mRNA/protein level in vivo.67,68 Zheng Y et al published a study focusing on cholesterol metabolism in osteoporosis in 2021, they found that geniposide can effectively ameliorate dexamethasone-induced cholesterol accumulation via activating the GLP-1R/ABCA1 axis in MC3T3-E1 cells.69 Geniposide has significant beneficial effects on cholesterol metabolism and lipid accumulation in HepG2 cells. After HepG2 cells were treated with geniposide, the expression of ABCA1 and AMPK mRNA significantly increased, while that of Cytochrome P450 Family 7 Subfamily A Member 1 (CYP7A1) and Liver X receptor alpha (LXRA) mRNA was significantly reduced.68 Geniposide and chlorogenic acid combination can significantly reduce hepatic TG, TC, and FFA via SCD-1 suppression in the liver.66
Regulating Lipid Transport
Lipid transportation in the blood depends on plasma lipoproteins. High-density lipoproteins (HDL) snatch the cholesterol from other lipoproteins or the peripheral tissues and bring it back to the liver, which is called reverse cholesterol transport (RCT). In the circulation, VLDL, produced in the liver, release TGs and FFA.71 Hepatic lipase removes TGs from intermediate-density lipoprotein, forming LDL. LDL has a high cholesterol content and it can be removed from circulation through binding to LDL receptors in extrahepatic tissues and the liver.72
Geniposide attenuates the development of atherosclerosis and reduces serum TC, TG, and LDL levels in ApoE-/- mice.70 In vitro and in vivo experiments have revealed that geniposide can modify the efflux-related proteins and lipoproteins to regulate cholesterol uptake, thus balancing the lipid transport levels and eventually inhibiting the formation of foam cells. These advantages seem to be mediated by the downregulation of P-p38 mitogen-activated protein kinase (MAPK) and P-AKT.
There are two main methods of assuaging hypercholesterolemia. The first is to increase the transformation of cholesterol into bile acids in the liver and the other is enhanced RCT, through which cholesterol in the plasma can be returned to the liver for catabolism. On the one hand, geniposide improves the hepatic synthesis of bile acids via passivating the negative feedback regulation of bile acids regulated by Farnesoid X receptor (FXR).73 On the other hand, geniposide facilitates the RCT, more cholesterol is directed from the circulation back to the liver, exerting the hypolipidemic effect of geniposide indirectly.73 Hwa-Young Lee74 displayed an unusual yet very clear design solution, their study was conducted using the Eucommia ulmoides Oliver extract, and its active constituents, aucubin and geniposide. They found that geniposide reduces hepatic lipid accumulation and secretion of apolipoprotein B, and they believe the mechanism may be linked to ER stress and hepatic dyslipidemia.
Discussion
Glucolipid metabolism is complex and variable, often involving multiple pathological reactions. Over the past few decades, a growing number of animal and cell model studies have revealed the potential of geniposide to ameliorate glucolipid metabolism disorders. We summarized basic information about various in vitro and in vivo studies on the pharmacological effects of geniposide on glucolipid metabolism in this review.
In terms of glucose metabolism, geniposide can increase glucose utilization and insulin production, protect pancreatic islet β-cells, inhibit IR and hepatic glucose production, and suppress gluconeogenesis. In the aspect of lipid metabolism, geniposide can promote lipid decomposition, inhibit lipogenesis, and regulate lipid transport. Geniposide regulates a wide range of factors related to glucose and lipid metabolism, including metabolic regulators AMPK, Glut2\Glut4\GLP-1R, which are indispensable in glucose metabolism, and PPARα/SREBP1c/ LXR/FXR, which regulate lipid metabolism. Such multi-pathway regulation improves the entire glycolipid metabolism network in a three-dimensional manner. However, there are still some challenges that need to be addressed.
First of all, the current research on geniposide is restricted to whole animal and cell experiments and has not included any clinical trials. Therefore, further clinical studies are essential to determine the role of geniposide in clinical therapy.
Secondly, thorough studies and precise targets are required. Studies on the effect of geniposide on glucolipid metabolism are sometimes unconvincing and have not been verified via rigorous experiments using knockout mice or cell lines. There is insufficient evidence supporting the results and conclusions of these studies. At present, research focusing on the pathological mechanisms of geniposide has not included a specific target, and future studies should be more accurate.
Furthermore, the efficacy and toxicity of geniposide need to be confirmed. We investigated several studies on geniposide toxicity that were published in 2021, and found that chronic oral toxicity in rats resulted in an impact on serum biochemical, urinary, and hematological parameters, and affected related organ weights. However, studies which reported that geniposide improves glucose and lipid metabolism have not investigated the potential toxicity of geniposide. Further studies are warranted to verify the feasibility of geniposide as a drug and develop a safe procedure for its administration. Its efficacy should also be compared with that of the current major drugs for metabolic diseases.
Conclusion
Geniposide ameliorates lipid and glucose metabolic disorders, improving the entire glycolipid metabolism network in a three-dimensional manner at the level of molecular mechanism (Figure 6). Accumulating studies, related to the effect of geniposide on glucose and lipid metabolism, have been performed. While full-scale progress has been made, the underlying mechanism of glucolipid metabolism has not yet been utterly elucidated. Future research is warranted to explain both long- and short-term effects of geniposide on glucose and lipid metabolism. Our review affords the first systematic summary of research examining the effect of geniposide on glucolipid metabolism, which will be beneficial in the development of metabolic diseases therapy in the future and in obtaining more reproducible and reliable data.
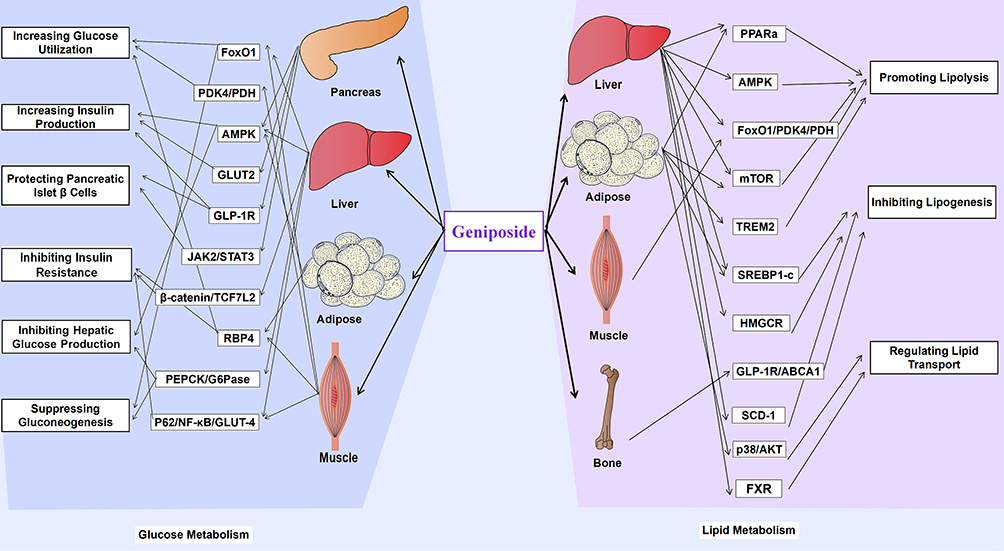 |
Figure 6 Molecular mechanisms involved in the regulation of glucolipid metabolism by geniposide. |
Abbreviations
NAFLD, non-alcohol fatty liver disease; GJE, Gardenia Jasminoides Ellis; EUO, Eucommia Ulmoides Oliv; RO, Rehmannia Officinalis; RS, Radix Scrophulariae; LPO, hepatic lipid peroxidation; GST, glutathione-S-transferase; GSH, glutathione; GPx, glutathione peroxidase; CuZn-SOD, copper- and zinc-containing superoxide dismutase; ATP, adenosine triphosphate; RBP4, retinol-binding protein 4; GSIS, Glucose-stimulated insulin secretion; GLP-1R, glucagon-like peptide 1 receptors; GLUT2, glucose transporter 2; AMPK, activated protein kinase; PDI, protein disulfide isomerase; ERO1, endoplasmic reticulum oxidoreductin 1; IAPP, amyloid polypeptide; Txnip, thioredoxin-interacting protein; ER, endoplasmic reticulum; IR, Insulin Resistance; RBP4, retinol-binding protein 4; TAGs, triacylglycerols; CEs, cholesterol esters; LDs, lipid droplets; ACaCa, acetyl-CoA carboxylase; FASN, fatty acid synthase; SFAs, saturated fatty acids; MUFAs, monounsaturated fatty acids; SCD1, stearoyl-CoA desaturase 1; chREBP, carbohydrate-responsive element-binding protein; LDL-c, low-density lipoprotein cholesterol; VLDL, very-low-density lipoproteins; HDL, High-density lipoproteins; RCT, reverse cholesterol transport.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Rana S, Ali S, Wani HA, et al. Metabolic syndrome and underlying genetic determinants-A systematic review. J Diabetes Metab Disord. 2022;21(1):1095–1104. doi:10.1007/s40200-022-01009-z
2. Ikegami H, Hiromine Y, Noso S. Insulin-dependent diabetes mellitus in older adults: current status and future prospects. Geriatr Gerontol Int. 2022;8:595–609.
3. Schrack JA, Dougherty RJ, Corkum A, et al. Impact of time in motion on blood pressure regulation among patients with metabolic syndrome. Curr Hypertens Rep. 2022. doi:10.1007/s11906-022-01202-8
4. Hasebe T, Hasebe N. Impact of risk factors related to metabolic syndrome on acute myocardial infarction in younger patients. Hypertens Res. 2022;45:287–299.
5. Rahman MM, Dhar PS. Exploring the plant-derived bioactive substances as antidiabetic agent: an extensive review. Bio Pharmacother. 2022;152:113–217. doi:10.1016/j.biopha.2022.113217
6. Tian J, Qin S, Han J, et al. A review of the ethnopharmacology, phytochemistry, pharmacology and toxicology of Fructus Gardeniae (Zhi-zi). J Ethnopharmacol. 2022;289:114984. doi:10.1016/j.jep.2022.114984
7. Shan M, Yu S, Yan H, et al. A review on the phytochemistry, pharmacology, pharmacokinetics and toxicology of geniposide, a natural product. Molecules. 2017;22(10):255–278. doi:10.3390/molecules22101689
8. Lv S, Ding Y, Zhao H, et al. Therapeutic potential and effective components of the Chinese herb gardeniae fructus in the treatment of senile disease. Aging Dis. 2018;9(6):1153–1164. doi:10.14336/AD.2018.0112
9. Fan X, Lin L, Cui B, et al. Therapeutic potential of genipin in various acute liver injury, fulminant hepatitis, NAFLD and other non-cancer liver diseases: more friend than foe. Pharmacol Res. 2020;159:104–145. doi:10.1016/j.phrs.2020.104945
10. Ran D, Hong W, Yan W, et al. Properties and molecular mechanisms underlying geniposide-mediated therapeutic effects in chronic inflammatory diseases. J Ethnopharmacol. 2021;273:113–158. doi:10.1016/j.jep.2021.113958
11. Liu L, Wu Q, Chen Y, et al. Updated pharmacological effects, molecular mechanisms, and therapeutic potential of natural product geniposide. Molecules. 2022;27(10):7900–7969.
12. Zhou YX, Zhang RQ, Rahman K, et al. Diverse pharmacological activities and potential medicinal benefits of geniposide. Evid Based Complement Alternat Med. 2019;2019:492. doi:10.1155/2019/4925682
13. Li N, Li L, Wu H, et al. Antioxidative property and molecular mechanisms underlying geniposide-mediated therapeutic effects in diabetes mellitus and cardiovascular disease. Oxid Med Cell Longev. 2019;2019:748.
14. Zhang X, Liu K, Shi M, et al. Therapeutic potential of catalpol and geniposide in Alzheimer’s and Parkinson’s diseases: a snapshot of their underlying mechanisms. Brain Res Bull. 2021;174:281–295. doi:10.1016/j.brainresbull.2021.06.020
15. Zhang W, Zhang F, Hu Q, et al. The emerging possibility of the use of geniposide in the treatment of cerebral diseases: a review. Chin Med. 2021;16(1):86–102. doi:10.1186/s13020-021-00486-3
16. Zhang A, Sun H, Yuan Y, et al. An in vivo analysis of the therapeutic and synergistic properties of Chinese medicinal formula Yin-Chen-Hao-Tang based on its active constituents. Fitoterapia. 2011;82(8):1160–1168. doi:10.1016/j.fitote.2011.07.014
17. Wu X, Zhou Y, Yin F, et al. Quality control and producing areas differentiation of Gardeniae Fructus for eight bioactive constituents by HPLC-DAD-ESI/MS. Phytomedicine. 2014;21(4):551–559. doi:10.1016/j.phymed.2013.10.002
18. Fu XM, Yang C, Wu ZG, et al. 不同采收期栀子中8个有效成分含量动态变化与颜色的相关性研究. [Correlation between color and content of eight components of Gardeniae Fructus at different harvest time]. Zhongguo Zhong Yao Za Zhi. 2020;45(13):3191–3202. Chinese. doi:10.19540/j.cnki.cjcmm.20200229.202
19. Chang R, Liu J, Luo Y, et al. Isoflavones’ effects on pharmacokinetic profiles of main iridoids from gardeniae fructus in rats. J Pharm Anal. 2020;10(6):571–580. doi:10.1016/j.jpha.2019.11.004
20. Wang Y, Feng F. Evaluation of the Hepatotoxicity of the Zhi-Zi-Hou-Po Decoction by Combining UPLC-Q-Exactive-MS-Based Metabolomics and HPLC-MS/MS-based geniposide tissue distribution. Molecules. 2019;24(3):123–145.
21. Jeong SH, Jang JH, Cho HY, et al. Simultaneous determination of three iridoid glycosides of Rehmannia glutinosa in rat biological samples using a validated hydrophilic interaction-UHPLC-MS/MS method in pharmacokinetic and in vitro studies. J Sep Sci. 2020;43(22):4148–4161. doi:10.1002/jssc.202000809
22. Yang X, Li J, Yang X, et al. Comparative pharmacokinetics of geniposidic acid, genipin-1-β-gentiobioside, geniposide, genipin, and crocetin in rats after oral administration of crude gardeniae fructus and its three processed products using LC-MS/MS. Evid Based Complement Alternat Med. 2020;2020:164–261.
23. Yang L, Bi L, Jin L, et al. Geniposide ameliorates liver fibrosis through reducing oxidative stress and inflammatory response, inhibiting apoptosis and modulating overall metabolism. Front Pharmacol. 2021;12:772–795.
24. Nguyen L, Ahn SH, Nguyen UT, et al. Geniposide, a principal component of gardeniae fructus, protects skin from diesel exhaust particulate matter-induced oxidative damage. Evid Based Complement Alternat Med. 2021;2021:884–958. doi:10.1155/2021/8847358
25. Shen Y, Wang X, Shen X, et al. Geniposide possesses the protective effect on myocardial injury by inhibiting oxidative stress and ferroptosis via activation of the Grsf1/GPx4 Axis. Front Pharmacol. 2022;13:879. doi:10.3389/fphar.2022.879870
26. Gheitasi I, Savari F, Akbari G, et al. Molecular mechanisms of hawthorn extracts in multiple organs disorders in underlying of diabetes: a review. Int J Endocrinol. 2022;2022:725–768. doi:10.1155/2022/2002768
27. Tuttle KR, Agarwal R, Alpers CE, et al. Molecular mechanisms and therapeutic targets for diabetic kidney disease. Kidney Int. 2022;102:248–260. doi:10.1016/j.kint.2022.05.012
28. Li Y, Hei H, Zhang S, et al. PGC-1α participates in tumor chemoresistance by regulating glucose metabolism and mitochondrial function. Mol Cell Biochem. 2022. doi:10.1007/s11010-022-04477-2
29. Sponton CH, de Lima-Junior JC, Leiria LO. What puts the heat on thermogenic fat: metabolism of fuel substrates. Trends Endocrinol Metab. 2022;1:965–996.
30. Li Y, Pan H, Zhang X, et al. Geniposide improves glucose homeostasis via regulating FoxO1/PDK4 in skeletal muscle. J Agric Food Chem. 2019;67(16):4483–4492. doi:10.1021/acs.jafc.9b00402
31. Liu J, Song C, Nie C, et al. A novel regulatory mechanism of geniposide for improving glucose homeostasis mediated by circulating RBP4. Phytomedicine. 2022;95:153–162. doi:10.1016/j.phymed.2021.153862
32. Ishihara H. Metabolism-secretion coupling in glucose-stimulated insulin secretion. Diabetol Int. 2022;13(3):463–470. doi:10.1007/s13340-022-00576-z
33. Guo LX, Xia ZN, Gao X, et al. Glucagon-like peptide 1 receptor plays a critical role in geniposide-regulated insulin secretion in INS-1 cells. Acta Pharmacol Sin. 2012;33(2):237–241. doi:10.1038/aps.2011.146
34. Liu J, Guo L, Yin F, et al. Geniposide regulates glucose-stimulated insulin secretion possibly through controlling glucose metabolism in INS-1 cells. PLoS One. 2013;8(10):e78315. doi:10.1371/journal.pone.0078315
35. Guo LX, Liu JH, Yin F. Regulation of insulin secretion by geniposide: possible involvement of phosphatidylinositol 3-phosphate kinase. Eur Rev Med Pharmacol Sci. 2014;18(9):1287–1294.
36. Jiang XQ, Shen SL, Li WZ, et al. 京尼平苷调节胰腺β细胞GLUT2糖基化的分子机制. [Molecular mechanism of geniposide in regulating GLUT2 glycosylation in pancreatic β cells]. Zhongguo Zhong Yao Za Zhi. 2021;46(14):3643–3649. Chinese. doi:10.19540/j.cnki.cjcmm.20200118.401
37. Hao Y, Liu C, Yin F, et al. 5’-AMP-activated protein kinase plays an essential role in geniposide-regulated glucose-stimulated insulin secretion in rat pancreatic INS-1 β cells. J Nat Med. 2017;71(1):123–130. doi:10.1007/s11418-016-1038-5
38. Liu C, Hao Y, Yin F, et al. Geniposide balances the redox signaling to mediate glucose-stimulated insulin secretion in pancreatic β-cells. Diabetes Metab Syndr Obes. 2020;13:509–520. doi:10.2147/DMSO.S240794
39. Zhang Y, Yin F, Liu J, et al. Geniposide protects pancreatic INS-1E β cells from hIAPP-induced cell damage: potential involvement of insulin degrading-enzyme. Cell Biol Int. 2015;39(4):373–378. doi:10.1002/cbin.10394
40. Liu C, Hao Y. Yin, et al. Geniposide protects pancreatic β cells from high glucose-mediated injury by activation of AMP-activated protein kinase. Cell Biol Int. 2017;41(5):544–554. doi:10.1002/cbin.10758
41. Liu CY, Hao YN, Yin F, et al. Geniposide accelerates proteasome degradation of Txnip to inhibit insulin secretion in pancreatic β-cells. J Endocrinol Invest. 2017;40(5):505–512. doi:10.1007/s40618-016-0591-9
42. Hao Y, Shen S, Yin F, et al. Unfolded protein response is involved in geniposide-regulating glucose-stimulated insulin secretion in INS-1 cells. Cell Biochem Funct. 2019;37(5):368–376. doi:10.1002/cbf.3414
43. Hjørne AP, Modvig IM, Holst JJ. The sensory mechanisms of nutrient-induced GLP-1 Secretion. Metabolites. 2022;12(5):234–263. doi:10.3390/metabo12050420
44. Liu J, Yin F, Xiao H, et al. Glucagon-like peptide 1 receptor plays an essential role in geniposide attenuating lipotoxicity-induced β-cell apoptosis. Toxicol In Vitro. 2012;26(7):1093–1097. doi:10.1016/j.tiv.2012.07.004
45. Cui LJ, Bai T, Zhi LP, et al. Analysis of long noncoding RNA-associated competing endogenous RNA network in glucagon-like peptide-1 receptor agonist-mediated protection in β cells. World J Diabetes. 2020;11(9):374–390. doi:10.4239/wjd.v11.i9.374
46. Yao DD, Yang L, Wang Y, et al. Geniposide promotes beta-cell regeneration and survival through regulating β-catenin/TCF7L2 pathway. Cell Death Dis. 2015;6(5):e1746. doi:10.1038/cddis.2015.107
47. Kojima K, Shimada T, Nagareda Y, et al. Preventive effect of geniposide on metabolic disease status in spontaneously obese type 2 diabetic mice and free fatty acid-treated HepG2 cells. Biol Pharm Bull. 2011;34(10):1613–1618. doi:10.1248/bpb.34.1613
48. Jiang H, Ma Y, Yan J, et al. Geniposide promotes autophagy to inhibit insulin resistance in HepG2 cells via P62/NF‑κB/GLUT‑4. Mol Med Rep. 2017;16(5):7237–7244. doi:10.3892/mmr.2017.7503
49. Zhao W, Pu M, Shen S, et al. Geniposide improves insulin resistance through AMPK-mediated Txnip protein degradation in 3T3-L1 adipocytes. Acta Biochim Biophys Sin (Shanghai). 2021;53(2):160–169. doi:10.1093/abbs/gmaa157
50. Guo L, Zheng X, Liu J, et al. Geniposide suppresses hepatic glucose production via AMPK in HepG2 Cells. Biol Pharm Bull. 2016;39(4):484–491. doi:10.1248/bpb.b15-00591
51. Yang SQ, Chen YD, Li H, et al. Geniposide and gentiopicroside suppress hepatic gluconeogenesis via regulation of AKT-FOXO1 Pathway. Arch Med Res. 2018;49(5):314–322. doi:10.1016/j.arcmed.2018.10.005
52. Wu SY, Wang GF, Liu ZQ, et al. Effect of geniposide, a hypoglycemic glucoside, on hepatic regulating enzymes in diabetic mice induced by a high-fat diet and streptozotocin. Acta Pharmacol Sin. 2009;30(2):202–208. doi:10.1038/aps.2008.17
53. Chew N, Ng CH, Truong E, et al. Non-alcoholic steatohepatitis drug development pipeline: an update. Semin Liver Dis. 2022;42:356–365.
54. Deng KQ, Huang X, Lei F, et al. Role of hepatic lipid species in the progression of nonalcoholic fatty liver disease. Am J Physiol Cell Physiol. 2022;23:244–298.
55. Rodrigues JE, Martinho A, Santa C, et al. Systematic review and meta-analysis of mass spectrometry proteomics applied to human peripheral fluids to assess potential biomarkers of schizophrenia. Int J Mol Sci. 2022;23(9):201–387.
56. Todisco S, Santarsiero A, Convertini P, et al. PPAR alpha as a metabolic modulator of the liver: role in the pathogenesis of nonalcoholic steatohepatitis (NASH). Biology. 2022;11:5.
57. Ma T, Huang C, Zong G, et al. Hepatoprotective effects of geniposide in a rat model of nonalcoholic steatohepatitis. J Pharm Pharmacol. 2011;63(4):587–593. doi:10.1111/j.2042-7158.2011.01256.x
58. Xu YL, Liu XY, Cheng SB, et al. Geniposide enhances macrophage autophagy through downregulation of trem2 in atherosclerosis. Am J Chin Med. 2020;48(8):1821–1840. doi:10.1142/S0192415X20500913
59. Fu C, Zhang X, Lu Y, et al. Geniposide inhibits NLRP3 inflammasome activation via autophagy in BV-2 microglial cells exposed to oxygen-glucose deprivation/reoxygenation. Int Immunopharmacol. 2020;84:106–147. doi:10.1016/j.intimp.2020.106547
60. Luo X, Wu S, Jiang Y, et al. Inhibition of autophagy by geniposide protects against myocardial ischemia/reperfusion injury. Int Immunopharmacol. 2020;85:106–159. doi:10.1016/j.intimp.2020.106609
61. Dusabimana T, Park EJ, Je J, et al. Geniposide improves diabetic nephropathy by enhancing ULK1-mediated autophagy and reducing oxidative stress through AMPK activation. Int J Mol Sci. 2021;22(4):109–124. doi:10.3390/ijms22041651
62. Liu DX, Zhang D, Hu WM, et al. Geniposide protection against Aβ(1-42) toxicity correlates with mTOR inhibition and enhancement of autophagy. J Integr Neurosci. 2021;20(1):67–75. doi:10.31083/j.jin.2021.01.242
63. Wang Y, Pan Y, Liu Y, et al. A new geniposidic acid derivative exerts antiaging effects through antioxidative stress and autophagy induction. Antioxidants. 2021;10(6):897–923. doi:10.3390/antiox10060897
64. Li Y, Zhang K, Liu J, et al. Geniposide suppresses thermogenesis via regulating PKA catalytic subunit in adipocytes. Toxicology. 2021;464:153–214. doi:10.1016/j.tox.2021.153014
65. Liang HQ, Lin MT, Zhao X, et al. 栀子苷改善大鼠非酒精性脂肪性肝病游离脂肪酸代谢的机制研究. [Mechanism of geniposide in improving free fatty acid metabolism in rats with non-alcoholic fatty liver disease]. Zhongguo Zhong Yao Za Zhi. 2016;41(3):470–475. Chinese. doi:10.4268/cjcmm20160319
66. Chen C, Xin X, Liu Q, et al. Geniposide and chlorogenic acid combination improves non-alcoholic fatty liver disease involving the potent suppression of elevated hepatic SCD-1. Front Pharmacol. 2021;12:653–661.
67. Shen B, Feng H, Cheng J, et al. Geniposide alleviates non-alcohol fatty liver disease via regulating Nrf2/AMPK/mTOR signalling pathways. J Cell Mol Med. 2020;24(9):5097–5108. doi:10.1111/jcmm.15139
68. Leng E, Xiao Y, Mo Z, et al. Synergistic effect of phytochemicals on cholesterol metabolism and lipid accumulation in HepG2 cells. BMC Complement Altern Med. 2018;18(1):122–167. doi:10.1186/s12906-018-2189-6
69. Zheng Y, Xiao Y, Zhang D, et al. Geniposide Ameliorated Dexamethasone-Induced Cholesterol Accumulation in Osteoblasts by Mediating the GLP-1R/ABCA1 Axis. Cells-Basel. 2021;10:12.
70. Shen D, Zhao D, Yang X, et al. Geniposide against atherosclerosis by inhibiting the formation of foam cell and lowering reverse lipid transport via p38/MAPK signaling pathways. Eur J Pharmacol. 2019;864:172–198. doi:10.1016/j.ejphar.2019.172728
71. Newman CB. Effects of endocrine disorders on lipids and lipoproteins. Best Pract Res Clin Endocrinol Metab. 2022;101–167. doi:10.1016/j.beem.2022.101667
72. Ribas-Latre A, Eckel-Mahan K. Nutrients and the circadian clock: a partnership controlling adipose tissue function and health. Nutrients. 2022;14(10):456–489. doi:10.3390/nu14102084
73. Liu J, Li Y, Sun C, et al. Geniposide reduces cholesterol accumulation and increases its excretion by regulating the FXR-mediated liver-gut crosstalk of bile acids. Pharmacol Res. 2020;152:104–131. doi:10.1016/j.phrs.2020.104631
74. Lee HY, Lee GH, Lee MR, et al. Eucommia ulmoides Oliver extract, aucubin, and geniposide enhance lysosomal activity to regulate ER stress and hepatic lipid accumulation. PLoS One. 2013;8(12):813–849. doi:10.1371/journal.pone.0081349
 © 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.