")
Back to Journals » Clinical Ophthalmology » Volume 18
Breaking-Down Barriers: Proposal of Using Cellulose Biosynthesis Inhibitors and Cellulase Enzyme as a Novel Treatment Modality for Vision Threatening Pythium Insidiosum Keratitis
Authors Gurnani B , Natarajan R, Mohan M, Kaur K
Received 3 December 2023
Accepted for publication 5 March 2024
Published 11 March 2024 Volume 2024:18 Pages 765—776
DOI https://doi.org/10.2147/OPTH.S450665
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Scott Fraser
Bharat Gurnani,1 Radhika Natarajan,2 Madhuvanthi Mohan,2 Kirandeep Kaur3
1Department of Cataract, Cornea and Refractive Surgery, ASG Eye Hospital, Jodhpur, Rajasthan, 342008, India; 2Department of Cornea and Refractive Surgery, Sankara Nethralaya Medical Research Foundation, Chennai, Tamil Nadu, 600006, India; 3Department of Pediatric Ophthalmology and Strabismus, ASG Eye Hospital, Jodhpur, Rajasthan, 342008, India
Correspondence: Bharat Gurnani, Department of Cornea and Refractive Surgery, ASG Eye Hospital, Jodhpur, Rajasthan, 342008, India, Tel +919080523059, Email [email protected]
Abstract: Pythium insidiosum, an Oomycete, causes severe keratitis that endangers vision. Its clinical, morphological, and microbiological characteristics are often indistinguishable from those of fungal keratitis, earning it the moniker “parafungus”. Distinctive clinical hallmarks that set it apart from other forms of keratitis include radial keratoneuritis, tentacles, marginal infiltration, and a propensity for rapid limbal spread. The therapeutic approach to Pythium keratitis (PK) has long been a subject of debate, and topical and systemic antifungals and antibacterials have been tried with limited success. Approximately 80% of these eyes undergo therapeutic keratoplasty to salvage the eye. Hence, there is a need to innovate for alternative and better medical therapy to safeguard these eyes. The resistance of Pythium to standard antifungal treatments can be attributed to the absence of ergosterol in its cell wall. Cell walls of plants and algae have cellulose as an essential constituent. Cellulose imparts strength and structure and acts as the “skeleton” of the plant. Fungal and animal cell walls typically lack cellulose. The cellular architecture of Pythium shares a similarity with plant and algal cells through the incorporation of cellulose within its cell wall structure. Inhibitors targeting cellulose biosynthesis (CBI), such as Indaziflam, Isoxaben, and Quinoxyphen, serve as critical tools for elucidating the pathways of cellulose synthesis. Furthermore, the enzymatic action of cellulase is instrumental for the extraction of proteins and DNA. To circumvent this issue, we hypothesize that CBI’s and cellulase enzymes can act on the Pythium cell wall and may effectively treat PK. The available literature supporting the hypothesis and proof of concept has also been discussed. We have also discussed these drugs’ molecular mechanism of action on the Pythium cell wall. We also aim to propose how these drugs can be procured and used as a potential medical management option for this devastating entity.
Keywords: Pythium insidiosum, keratitis, cellulose biosynthesis inhibitors, cellulase enzyme, antifungals
Introduction
Pythium insidiosum keratitis represents a recently identified ophthalmic concern characterized by its capacity to induce severe, vision-compromising infection of the cornea. This oomycete organism bears a striking resemblance to fungal pathogens in clinical presentation.1 Pythium species, particularly identified as “parafungus”, are classified within the Phylum Oomycota, belonging to the Kingdom Straminipila, the Class Oomycetes, Order Pythiales, and the Family Pythiaceae. The first Pythium insidiosum keratitis (PIK) case was documented in Thailand in 1988.1,2 Since its initial identification, Pythium insidiosum has proven to be a formidable mimic of fungal keratitis in clinical, morphological, and microbiological aspects, posing significant diagnostic and therapeutic challenges.2 PIK is clinically distinguished by a constellation of symptoms, including reticular dot infiltrates, tentacle-like projections, peripheral guttering accompanied by thinning of the corneal tissue, and rapid advancement to the limbal region.3 The management of this form of keratitis remains challenging due to its infrequent clinical manifestation, the paucity of specific diagnostic tools, its inherently aggressive nature, swift progression, typically unfavorable outcomes, and a propensity for recurrent episodes. PIK is uncommon and can result in vision-threatening sequelae.4 On smear analysis with 10% KOH, Gram stain, acridine orange, IKI-H2SO4, KOH, and lactophenol blue mount, there is the presence of septate or aseptate perpendicular hyphae, which can be of variable size.3,4 The smear characteristics closely resemble fungal keratitis, making it a diagnostic dilemma for many clinicians. When cultivated on media enriched with 5% sheep blood agar, chocolate agar, or other nutrient-rich agars, Pythium insidiosum gives rise to colonies that are characteristically dull grey to brown with a refractile appearance.5 Laboratory confirmation of Pythium insidiosum keratitis hinges on identifying zoospores using the leaf incarnation technique. The pharmacological approach to managing this condition, as outlined in scientific texts, includes the use of agents with antifungal and antibacterial properties that are efficacious against Pythium species.1,3 Microscopic analysis of Pythium’s cellular wall has demonstrated an absence of ergosterol, a component typically targeted by standard antifungal treatments. This deficiency renders the organism resistant to such interventions, exacerbating the risk of ocular complications.6 In one of the previous studies, in vivo analysis of samples reveals that the minimum inhibitory concentration (MIC) of antibacterials was greater than antifungals for PIK, which makes antibacterials the drug of choice.7 Pythium is a highly virulent, rapidly progressive keratitis requiring an early therapeutic keratoplasty with at least a 1–1.5 mm clearance margin. As per previous studies, approximately 70% of patients require surgical intervention, and a minority resolve with maximal medical therapy.8 In contrast to other keratitis, PIK has a poor prognosis with medical therapy. There is a need to develop, design, and discover new drug therapies to circumvent this challenge to combat this vision-threatening keratitis.9 It is well known that plant and algal cell walls have cellulose as the major component.10 Pythium cell walls also contain cellulose in the cell wall.11 Cellulose biosynthesis inhibitors (CBIs) and cellulose have a broad herbicidal action that targets cellulose in the cell wall.12 We hypothesize CBIs and cellulase can act on the Pythium cell wall and serve as a novel treatment modality in the future for the management of PIK. The available literature supporting the hypothesis and proof of concept has also been discussed. We have also discussed the molecular mechanism of action of these drugs on the Pythium cell wall. We also aim to propose how these drugs can be procured and can be used as a potential medical management option for this devastating entity.
Hypothesis
An Overview of Pythium Keratitis
Oomycetes represent a unique clade of eukaryotic organisms with fungoid characteristics. Initial classification aligned them with fungi, mirroring their morphological traits and ecological roles. Subsequent molecular and phylogenetic analyses have discerned substantial disparities between oomycetes and true fungi, leading to their reclassification under the stramenopiles.13 Oomycetes are characterized by their filamentous structure and heterotrophic growth mechanisms, capable of reproducing via both sexual and asexual means.1,2 Pythium species, categorized within the Oomycota phylum and the Oomycetes class under the Pythiaceae family, are commonly found in aqueous and terrestrial environments, predominantly within tropical and subtropical regions.2,3 Infections caused by Pythium can present in various forms, including cutaneous/subcutaneous, ocular, vascular, and disseminated types.2 The commonality in branching patterns, specifically the zygomaticus type, is a shared characteristic between pythium and various fungi. However, a distinct difference is observed in the composition of the pythium cell wall, which notably lacks ergosterol, a feature typically found in fungi.4,14 Additionally, it’s not just P. insidiosum that’s implicated in oomycete-related ocular infections. Species such as Lagenidium spp. and P. aphanidermatum are also recognized as causative agents. A notable case involved a patient from Thailand suffering from severe keratitis, which proved unresponsive to standard antibacterial and antifungal treatments, where Lagenidium sp. was identified as the pathogen”.15
Pythium keratitis is caused by an aquatic oomycete called Pythium insidiosum.13 It manifests as an aggressive keratitis due to their highly virulent nature, rapid progression, poor response to conventional antifungal therapy, and high recurrence rate.2 Previous encounters with environments comprising tainted aquatic sources or decaying plant material have been identified as considerable contributory elements for the onset of this infection.4 Despite their morphological similarities to fungi, these organisms exhibit resistance to antifungal agents, a trait attributed to the absence of ergosterol in their cellular walls. When the infective zoospores in the pythium come into contact with the ocular surface in a damp environment, they secrete glycoproteins, increasing the adhesion of zoospores to the epithelium.2,3,6 Incubation at physiological body temperatures facilitates the maturation of zoospores into hyphal forms, which then invade the host’s vascular tissues, thereby enhancing their ability to permeate corporeal tissues. From a therapeutic standpoint, such infections exhibit a more favorable response to antibacterial agents, including linezolid and azithromycin.7 Nevertheless, given the aggressive nature of the disease’s advancement, prompt intervention with penetrating keratoplasty, ensuring a minimum of 1mm of uninfected margins, remains the cornerstone of clinical management.2
Pythium is commonly confused with fungal keratitis due to similar zygomaticus branching patterns. But unlike fungal cell walls, which contain cellulose, beta-glucan, chitin, and, more importantly, ergosterol, the cell wall of pythium lacks chitin and ergosterol. This renders the common antifungal medications ineffective in the treatment of pythium keratitis.16
Clinical Features
Differentiating Pythium keratitis from fungal keratitis can present challenges; however, distinctive clinical manifestations such as stromal infiltrates exhibiting a net-like configuration, tentacular projection at the periphery of the ulcer, and circumferential grooving tend to be indicative of Pythium keratitis.1,2 Additionally, the propensity for Pythium infections to rapidly extend towards the limbus and cause marginal thinning accompanied by gutter formation further aids in its clinical identification.17
Microbiological Findings
On KOH wet mount, Pythium is seen as sparsely septate hyphae, which may mimic the slender septate or aseptate hyphae with perpendicular branches of fungi. This makes it quite challenging to diagnose on wet mount alone.2 The filaments of fungi stain with Periodic Acid Schiff stain as bright magenta, but there is a difference in staining in Pythium. The morphological presentation of these hyphae ranges from shades of pink to pale magenta; occasionally, they may not absorb stain. This staining variability stems from the presence of cellulose in the cell walls of Pythium, which interacts less robustly with acid-based stains due to its pectin and chitin composition.18 Differentiating Pythium insidiosum hyphae from those of hyaline fungi under direct microscopy remains a complex task due to these subtle staining characteristics. Definitive diagnosis is with the help of microbiological culture, but these are difficult due to the low zoospore formation rate.19
Characteristics of Cell Wall Architecture and Its Comparison with Cell Wall of Pythium
General Architecture of a Cellulitic Cell Wall
The fundamental architecture of the cell wall is constituted by a complex array of polysaccharides, protein, and aromatic compounds matrix. These components’ biochemical changes, interaction, and rearrangement provide a rigid and flexible cell wall architecture.20 The mechanical integrity and robustness of plant cell walls are attributed to an intricate matrix of cellulose microfibrils, which are composed of extended linear chains of β-1,4-linked glucose units. This cellulose network not only imparts substantial tensile strength, due to its polysaccharide nature, but also plays a critical role in reinforcing the cell wall’s structure. The spatial organization of these cellulose microfibrils is closely correlated with the cell’s developmental stage.21 For instance, during phases of swift cellular expansion, microfibrils tend to be oriented perpendicularly to the axis of growth. Conversely, perturbations in cellulose synthesis or its microfibrillar organization can result in compromised cell integrity, often manifesting as cells that expand excessively and rapidly, leading to stunted growth in the plant’s organs.
Cell Wall Architecture of Pythium Insidiosum Keratitis
To understand the molecular mechanism of various drugs, we must know the Pythium insidiosum’s cell wall. The composition of the cell wall in Oomycetes, such as the genus Pythium, has yet to be fully elucidated.22 Contemporary research has delved into the cell wall’s makeup, revealing that unlike fungal cell walls, which are characterized by the presence of ergosterols, Oomycete cell walls are principally constituted of beta-glucans and cellulose. Investigations into Pythium spp. have identified the presence of a variety of glucan linkages. For instance, Pythium acanthicum has been observed to contain an amalgamation of β-(1,3)-, (1,6)-, and β-(1,4)-linked glucans within its hyphal walls.23 Further scrutiny into Pythium debaryanum has expounded upon the ultrastructural complexity and the polysaccharide components that forge its cell wall. Diverse treatment modalities, including acid and alkali treatments, application of Schweitzer’s reagent, and enzymatic digestion with β-1,3-glucanase, coupled with electron microscopic analysis, have demonstrated that the acid-treated fraction is predominantly comprised of β-1,3-linked branched glucan. Conversely, the alkali-treated fraction is distinguished by a mix of 1.3-, 1.6-, and 1,3,6-linked glucans, suggesting a highly branched duality. The fraction processed with Schweitzer’s reagent revealed β-1,4-linked glucan. In terms of structural architecture, the cell wall is constructed of cellulosic microfibrils arranged into a delicate network that is interspersed with β-1,3-glucan including β-1,6-linkages. These cellulosic microfibrils are embedded within a matrix composed of an amorphous mixture of β-1,3-linked and β-1,6-linked branching glucans.24 Keeping in mind the cellulose-based architecture of the cell wall, we enlist the mechanism for various drugs which can be a potential treatment option for Pythium insidiosum keratitis in the future.
Possible Testing in vitro in Pythium Culture Plates
Natarajan et al examined the inhibitory potential of n-butyl-2-cyanoacrylate adhesive on cultures of Pythium insidiosum, which were isolated from corneal scrapings in a controlled in vitro environment”. They inoculated the isolates onto incubated culture plates, and cyanoacrylate glue was instilled. The sensitivity of Pythium species to the adhesive was demonstrated by the presence of a distinct zone indicating inhibition, with all four Pythium isolates exhibiting an unambiguous inhibitory zone.25
From a detailed literature review and from our clinical experience, we hypothesize that enzyme cellulase and Cellulose Biosynthesis Inhibitors (CBI) inhibitors have the potential to inhibit and destroy the cell wall of Pythium insidiosum. These two entities can act as a potential future treatment for PIK. The authors have discussed the supportive literature evidence and the mechanism of action of drugs inhibition cellulose. The authors have also enlisted the various CBIs which can act as potential drugs for PIK. Lastly, the authors have also discussed the future directions for clinical applications.
Evaluation of Hypotheses
Proposed Mechanism of Action of Drugs on Pythium Cell Wall
Cellulose Biosynthesis Inhibitors (CBIs)
Cellulose Biosynthesis Inhibitors constitute a category of compounds recognized as Group L herbicides by the Herbicide Resistance Action Committee (HRAC). These substances specifically target and impede the biosynthetic pathways responsible for cellulose production. CBIs are identified as chemical agents that disrupt the enzymatic pathways involved in the formation of cellulose. These inhibitors are further categorized according to their structural chemical characteristics.21 CBIs are used in the agricultural industry for weed control as cellulose killer herbicides in vineyards, grounds, orchids, lawns, and golf courses. CBI works by three possible modes of action for cellulose inhibition. First, the treated tissue shows reduced growth and alleviates expansion in a radial manner.26 Secondly, the synthesis of cellulose is observed to decrease in correlation with the dosage of the applied inhibitor, presenting a dose-dependent dynamic. Additionally, a swift inhibitory response is typically noted, often within a two-hour period post-integration of C-glucose into the cell wall’s cellulose structure. The intricacies within the cellulose biosynthetic pathways pose a challenge to fully elucidating the comprehensive inhibitory mechanisms of Cellulose Biosynthesis Inhibitors (CBIs). Another documented mode of action for CBIs involves the impediment of functional activity in the procuste-cesa6 mutant, which features a translational fusion of Yellow Fluorescent Protein (YFP) with Cellulose Synthase A6 (CESA6) under the control of its endogenous promoter. Laser scanning and confocal imaging can also help assess the CESA behaviour in live cells. CBI induces a change in the CESA particles at the plasma membrane level, cortical apparatus, and Golgi body complex. The three different proposed classifications for CBI based on mechanism of action are (1) First is drugs that cause clearance of CESA particles from the plasma membrane (2) Increased accumulation of CESA in the plasma membrane followed by slow or reduced movement of CESA in PM and (3) last is the changed CESA trajectory to PM and CESA speed at the PM focal plane. The important CBI are Cestrin, Indaziflam, and Acetobixan.21
Newer Cellulose Biosynthesis Inhibitors
Indaziflam
Indaziflam, an alkylazine herbicide, is key in developing Cellulose Biosynthesis Inhibitors (CBIs) and is associated with compounds like triazolam and AE F150944. Studies show indaziflam causes fluorescently-tagged cellulose synthase (CESA) complexes to accumulate at the plasma membrane, which move slower compared to untreated controls.27 This is thought to result from CESA aggregation after indaziflam exposure, a process that interestingly occurs without involvement from cellulose synthase interactive protein 1 (CSI1). However, the role of CSI3 is still unclear. In contrast, AE F150944 triggers different effects, specifically dispersing CESA complexes from the plasma membrane’s focal plane, leaving microtubules unaffected.28 Microscopic analysis reveals a scant presence of cellulase synthetase complex (CSC) at the plasma membrane. For Triazolam, there is no available freeze-fracture TEM imagery.29
Cestrin
CESTRIN, part of the CBI family, inhibits CESA trafficking, leading to blocked CESA exocytosis and lowered CSC levels on the plasma membrane. It increases CESA within SYP61 compartments, with SYP61 being essential for vesicle transport from the Golgi to the membrane. Proteomic studies have shown CESTRIN’s binding affinity with internal CSCs. Additionally, CESTRIN restricts CSC mobility on the plasma membrane, similar to how isoxaben and other CBIs limit CSC clearance at the membrane.30
Acetobixan
Acetobixan is derived from bacterial secretions and was discovered using subjective metabolic fingerprinting. Acetobixan has been observed to promote the dissociation of YFP-tagged CESA6 complexes from the plasma membrane. However, few compounds which cause resistance to quinoxphen or isoxaben were not cross resistance to acetobixan. This shows that these compounds can potentially disrupt the cellulose biosynthesis process. Based on a detailed literature review, we enlist various drugs which act as CBI and can be future management options for PIK by inhibiting the cell wall cellulose31 (Figure 1 and Table 1)
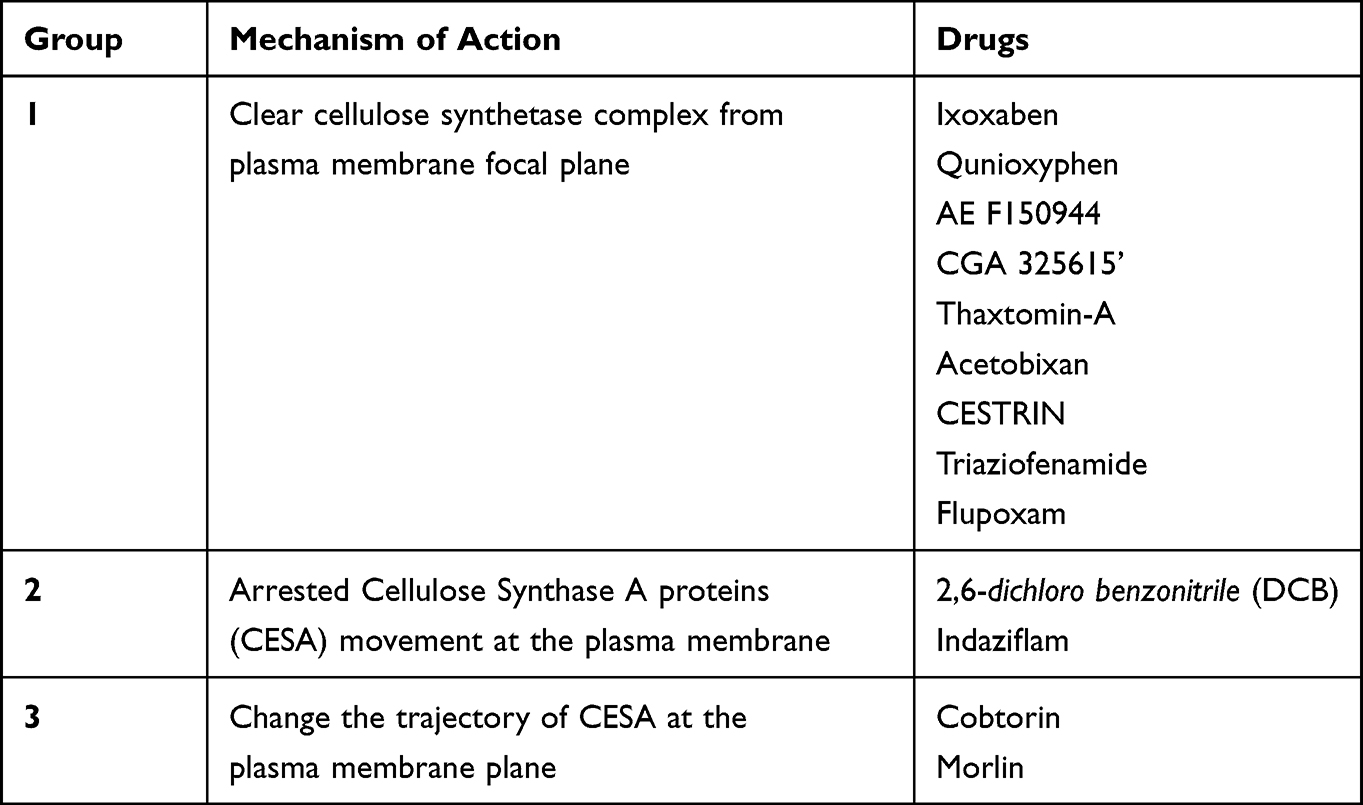 |
Table 1 Drug Classification of Cellulose Biosynthesis Inhibitors |
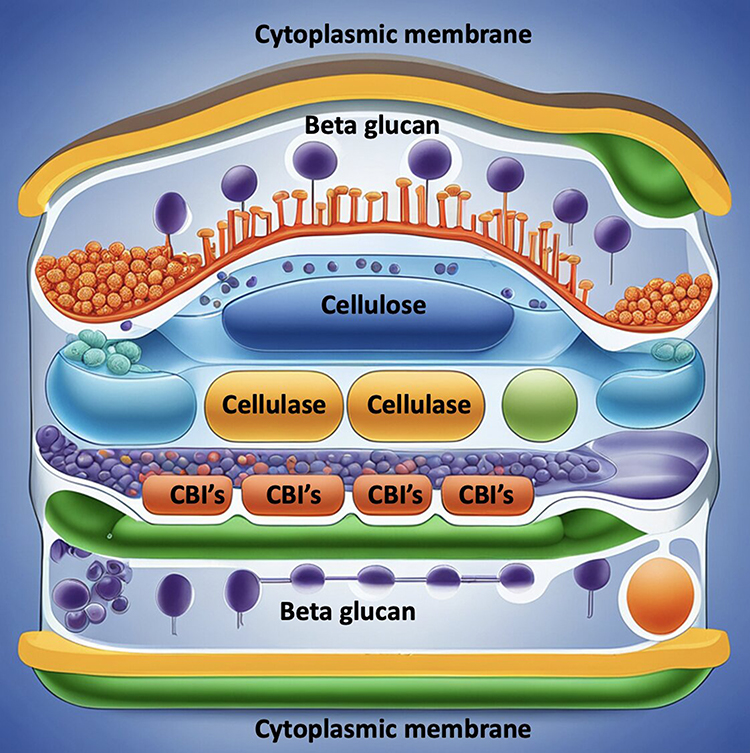 |
Figure 1 Depicts the schematic diagram of cell wall architecture of Pythium insidiosum keratitis and drugs such as Cellulose Biosynthesis Inhibitors (CBIs) and Cellulase acting on the cell wall. |
How to Destroy Cellulose/ Uses of Cellulase in Other Industries
Cellulose, the main structural component of plant cell walls, is a complex carbohydrate formed by the polymerization of glucose subunits into long, insoluble chains. Due to their tightly-ordered structure, cellulose chains resist hydrolytic breakdown or solubilization, posing a considerable challenge as they do not dissolve in water and require very high temperatures and pressures to break down.32 There are two primary methods for hydrolyzing cellulose. The chemical method involves using strong acids under extreme conditions, while the enzymatic method employs cellulase enzymes secreted by bacteria, which is considered the preferred method due to its relative non-toxicity.33 These cellulase enzymes fall into three classes: endo-(1,4)-β-D-glucanase, which cleaves internal bonds in the cellulose chain; exo-(1,4)-β-D-glucanase or cellobiohydrolase (CBH), acting at the chain ends to release disaccharides like cellobiose; and β-glucosidases that hydrolyze cellobiose into glucose. Their synergistic action is essential for complete cellulose degradation.33
Cellulases are a diverse group of enzymes produced by fungi and bacteria with significant roles across various sectors, such as food production, animal nutrition, viniculture, agronomy, biomass bio-refinement, and the manufacturing of paper, textiles, and detergents.34 In food technology, cellulase facilitates advanced extraction processes for fruits and vegetables, improving texture and stability while reducing nectar and puree viscosity, thereby enhancing yield without extra financial input.35 In animal nutrition, cellulase treatment improves the quality and nutritional value of silage and grain feed, and it may even eliminate toxins, leading to better digestion and nutrient absorption in animals.36
In the bioethanol sector, cellulase helps break down lignocellulosic biomass like sugarcane residues, corncobs, and forest byproducts into sugars for biofuel production.37 The pulp and paper industry has seen a significant decrease in energy consumption due to cellulase in biomechanical pulping, enhancing the physical characteristics of the pulp.36 In the textile industry, cellulase is pivotal in biostoning denim and improving the texture and color vibrancy of fabrics.38 Cellulase-containing detergents have become popular in home care for their effectiveness in brightening colors, softening fabrics, and removing soil from laundry.36 The enzyme’s application extends to olive oil extraction, enhancing yield and quality, waste management through the breakdown of cellulosic waste, and carotenoid extraction, which is critical in the production of natural pigments.36
While cellulase’s role in these sectors is well established, ongoing research and development continue to explore its potential in new applications. For instance, emerging studies suggest cellulase could play a role in enhancing the fermentability of agricultural residues, thereby contributing to more sustainable practices in biofuel production.37 It also shows promise in the development of novel waste treatment solutions, where its ability to decompose plant matter can be harnessed for more efficient recycling and reduction of landfill waste.36 Cellulase’s impact on industry and the environment underscores the importance of continued investment in biotechnology research. Advancements in enzyme technology may lead to more cost-effective and eco-friendly industrial processes, further broadening the applications of cellulase in various sectors and contributing to sustainability efforts globally.34–39
Methods- Empirical Data
Methodology for Testing Efficacy of Cellulase Enzyme in Pythium Colonies
To evaluate our theory, we will expose the ATCC strain of Pythium insidiosum, along with clinical variants, to assorted doses of cellulase. We will employ the agar dilution technique to ascertain the cellulase’s minimum inhibitory concentration (MIC) that arrests the organism’s growth. This method entails integrating progressive concentrations of the cellulase into a liquefied agar base, typically through serial binary dilutions, and then seeding a measured quantity of the microbe onto the agar’s surface. The MIC is established as the least amount of cellulase necessary to halt microbial proliferation during optimal growth conditions. This assay is applicable for gauging the sensitivity of bacterial and fungal pathogens. The agar dilution is particularly advantageous when multiple strains are assessed against a single agent or when the tested substance obscures growth visibility in a liquid medium due to its pigment. Historically, this approach has been a preferred method for determining the efficacy of antifungal treatments against organisms such as Candida, Aspergillus, Fusarium, and dermatophytes.40 Two separate investigations have been conducted utilizing the agar dilution assay to assess the in vitro efficacy of n-butyl cyanoacrylate adhesive against fungal pathogens and Pythium species. In the initial study, Dogan and the team probed the antifungal potential of n-butyl-2-cyanoacrylate on various fungal species.41 Concurrently, Natarajan and collaborators explored the suppressive impact of the same compound on isolates of Pythium insidiosum.25 In their investigation, Dogan et al procured clinical specimens from an array of fungal infections, encompassing corneal and skin abscesses, skin ulcers, and sputum samples.41 In the subsequent research, Pythium strains were collected from individuals diagnosed with infectious keratitis, utilizing corneal scraping techniques for sample acquisition.
Preparation & Inoculation
Fungal colonies were prepared in saline to the turbidity of the 0.5 McFarland standard, setting yeast and molds at 1.5 x 10^8 CFU/mL, with Fusarium species denser at 3 x 10^8 CFU/mL. These suspensions were uniformly applied to RPMI 1640 agar plates using a swabbing technique, followed by incubation at 35°C. To test microbial susceptibility, n-butyl-2-cyanoacrylate (nB2CA) was applied to the center of each agar plate, forming 7 mm disks. Post-incubation, a clear zone around the disks indicated susceptibility as per the Clinical Microbiology Procedures Handbook, while no zone suggested resistance. Inhibition zones were sampled, re-cultured, and analyzed to confirm the results.40,42,43 Separately, for Pythium insidiosum and other fungi, spores were suspended and spread on agar plates, then tested with a 6 mm nB2CA drop. The plates were incubated at 25°C in a Bio-Oxygen Demand (BOD) incubator. After 48 hours, the presence or absence of an inhibition zone was recorded to determine nB2CA’s antifungal efficacy. Fungal identification utilized morphological assessment and PCR targeting the ITS region. Swabs from inhibition zones were reinoculated to verify susceptibility.41 The studies illustrated nB2CA’s inhibitory effects against fungal and Pythium isolates. Additionally, the application of nB2CA in these studies is significant in understanding its potential as an antifungal agent. Its ability to create clear inhibition zones indicates a strong antifungal activity, making it a promising candidate for treating fungal infections. The consistent use of PCR for genetic identification ensures accurate species recognition, crucial for targeting specific fungal pathogens. The dual approach of the studies, combining physical growth inhibition with molecular techniques, provides a robust framework for future antifungal research. The re-culturing of inhibition zones serves as a critical step in confirming the initial findings and ensuring the reliability of nB2CA’s antifungal properties.40–43
Relevance of Hypothesis and Discussion
Pythium insidiosum keratitis is an aggressive infection that poses substantial management difficulties. Traditional treatments like antibacterials and antifungals are often inadequate. This highlights the urgent need for innovative treatments, specifically those targeting the unique cell wall composition of Pythium. Enzymes such as CBI and cellulase could provide a solution by disrupting the cell wall, curbing microbial growth early in the infection, and potentially improving the long-term prognosis of PIK.1
Cellulases are enzymes within the glycoside hydrolase family that catalyze the decomposition of cellulose into sugars by acid-base catalysis, a process that involves essential nucleophile and electrophile components.44 The reaction can change the sugar’s configuration, depending on the enzyme’s specific active site orientation. These enzymes are produced by a variety of microorganisms found in diverse environments like soil and decomposing organic matter. Notably, microbes such as Clostridium and Aspergillus are recognized for their potent cellulase activity, which is critical in various applications due to the enzymes’ intricate structure comprising functional domains and modules. Cellulases are synthesized in the presence of cellulose, highlighting their adaptive response to environmental cues and their significance in natural and industrial processes.45,46
In the structure of fungal cellulases, two distinct domains are typically present: the catalytic domain (CD), which facilitates the hydrolytic reaction, and the cellulose-binding module (CBM), tethered to the CD by a short polypeptide linker at the N-terminus, which often contains an abundance of serine and threonine residues and spans around 35 amino acids. Cellulosomes, multi-enzyme complexes, differ from solitary cellulase enzymes in their architecture, featuring cohesin modules for scaffolding and dockerin modules for enzymatic binding. Free cellulases, on the other hand, possess CBMs that anchor them to cellulose, whereas within a cellulosomal assembly, dockerins fulfill this role, with a singular scaffold-bound CBM directing the complex to the cellulose substrate.46
In the quest for diverse cellulolytic and hemicellulolytic enzymes, fungi and bacteria have been extensively utilized, with an emphasis traditionally placed on fungi for their robust secretion of uncomplicated cellulases and hemicellulases. Bacteria, however, are becoming increasingly recognized for their rapid proliferation and the ability to produce intricate multi-enzyme systems, thriving in a multitude of ecological settings. Their resilience in adverse conditions allows them to generate stable enzymes that enhance the efficiency of bioconversion. Particularly, the cell membranes of thermophilic bacteria, enriched with saturated fatty acids, confer a hydrophobic barrier that preserves cellular structure under elevated thermal conditions.47
Enhancements in the commercial production of cellulases are often achieved through genetic advancements and the application of mutagenic techniques to parental strains. Optimizing strains is a strategic method to augment cellulase output. While wild-type strains exhibit unique characteristics suitable for industrial use, refinement of these strains can further harness their innate capacities by altering or removing specific traits. Techniques such as recombinant DNA technology, site-directed mutagenesis, protoplast fusion, and random mutagenesis play pivotal roles in this enhancement. Additionally, immobilizing cellulase enzymes onto solid supports can significantly increase the efficiency of their catalytic function for industrial purposes.47
The fungus Hypocrea jecorina is known for synthesizing a comprehensive suite of cellulases, categorized into cellobiohydrolases (CBH), endoglucanases (EG), and β-glucosidases. This organism has revealed the existence of genes coding for two CBHs, cbh1 and cbh2; four EGs, egl1, egl2, egl3, and egl5; and a single β-glucosidase. The regulatory sequences of the cbh1, cbh2, egl1, and egl2 genes feature CRE1 binding sites, illustrating a sophisticated regulatory mechanism through carbon catabolic repression, which modulates these genes’ expression.48
In Hypocrea jecorina, the Activator of Cellulase Expression II (ACEII) protein interacts with the promoter regions of cellulase genes such as cbh1, cbh2, egl1, and egl2, playing a crucial role in regulating their expression.46 Conversely, the Activator of Cellulase Expression I (ACEI) gene, despite its presence on cbh1 promoters, functions as an inhibitor of cellulase gene expression.49 The process known as glucose repression of cellulase is thought to be orchestrated by the carbon catabolic repressor protein CRE1 in this organism.50 The cellulase promoters contain not only binding sites for CRE1 but also for transcriptional activators like Activator of Cellulase Expression II (ACEII) along with CCAAT sequences, which are known to bind to general transcriptional activator complexes, commonly referred to as HAP proteins.51 Suto and Tomita have offered an in-depth examination of cellulase gene expression, revealing that a sensor enzyme is constantly active, breaking down cellulose into oligosaccharides. These oligosaccharides subsequently enter the bacterial cell and trigger the activation of cellulase gene expression.52
Anaerobic protists that break down cellulose are predominantly found in the rumen, where they aid in decomposing plant matter. In the hindgut of lower termites, protists with flagella play a primary role in cellulose degradation. Similarly, wood-consuming cockroaches have a range of symbiotic protists in their hindgut that help them digest diets rich in cellulose. Protists often take in cellulose particles into their food vacuoles, where they break them down using enzymes known as cellulases. Despite this, there’s limited knowledge about the molecular structure of cellulases from protists. This is largely because cultivating the anaerobic community of gut protists poses challenges. Although only a few species have been grown in isolation, recent advancements in culture-free methods have aided in identifying and studying cellulases from these protists. Through PCR techniques, we obtained and sequenced fifteen complete cDNA clones from the protist population in the termite species, The termite Reticulitermes speratus has been studied for its cellulase enzymes, which have shown remarkable homology to a set of 45 cellulases identified in hypermastigote protists. These protist-derived cellulolytic enzymes are unique in that they consist solely of catalytic domains, lacking accessory modules such as carbohydrate-binding modules (CBD). The process of cellulose degradation in these organisms involves the physical breakdown of ingested cellulosic material by the host to increase the surface area available for enzyme action. Following this mechanical processing, the cellulosic substrates are endocytosed into food vacuoles for further enzymatic degradation.
Although these protist cellulases do not contain CBDs — typically responsible for tethering the enzyme to its substrate and concentrating the enzyme at the polysaccharide surface — the high concentration of enzymes and the expanded surface area of processed cellulose within the food vacuoles are thought to compensate for this absence.
In the case of the wood-feeding termite Coptotermes formosanus, a trio of symbiotic parabasalian flagellates — namely Spirotrichonympha leidyi, Holomastigotoides mirabile, and Pseudotrichonympha grassii — are integral to the digestion of its cellulose-rich diet. Notably, P. grassii is adapted to process high molecular weight cellulose particles, while H. mirabile and S. leidyi are more inclined to break down smaller, low molecular weight cello-oligosaccharides.53
Assessment of Toxicity
A study conducted by the EFSA Food Panel evaluated cellulase based on its ability to hydrolyze carboxymethyl cellulose (CMC), with the reaction conditions set at pH 4.8 and a temperature of 50°C over a 10-minute duration. Cellulase’s enzymatic function is assessed by its capacity to liberate reducing carbohydrates that, upon reacting with 3.5-dinitrosalicylic acid (DNS), yield a colored product. The enzyme’s activity is gauged in comparison to a standard and is denoted in carboxymethyl cellulose units per gram (CMCU/g). A single unit of cellulase activity equates to the enzyme quantity required to produce 1 micromole of glucose reducing sugar equivalents each minute within the test’s parameters.
The food enzyme’s operational characteristics were profiled, revealing an optimal temperature of approximately 65°C at a pH of 4.5 and an optimal pH range of 4.5–5.5 at 50°C. Its thermostability was examined after a 30-minute pre-incubation at various temperatures, maintaining a pH of 4.5. The enzyme’s activity was found to diminish significantly above 55°C and was entirely inactive when exposed to 75°C or higher. In an extended 90-day oral toxicity assay, the panel discerned a no observed adverse effect level (NOAEL) of 97.6 mg of total organic solids (TOS) per kilogram of body weight per day, which was the highest dosage evaluated.54
Purification
Due to their broad range of industrial applications, cellulases require purification and characterization to eliminate impurities and optimize their functionality for specific tasks. Elshafei et al detailed a purification process for endoglucanase from Aspergillus terreus using a combination of acetone fractionation and a Sepharose-4B gel filtration chromatography. Nehad et al have emphasized that to elucidate the mechanism of cellulose hydrolysis, cellulases must be purified and characterized, employing various isolation techniques. The raw enzyme extract may undergo initial purification via ammonium sulfate precipitation and subsequently undergo refinement through methods such as ion-exchange chromatography and high-performance liquid chromatography (HPLC).
Nguyen and Quyen described a purification regimen for endoglucanase from Aspergillus oryzae, beginning with 65% ammonium sulfate precipitation, followed by dialysis, and culminating with a Sephadex G-100 gel filtration column balanced with a 50 mM potassium phosphate buffer at pH 7.5. Similarly, the β-glucosidase produced by fusants of A. oryzae and A. niger was purified using ammonium sulfate precipitation, ion-exchange, and gel filtration chromatography with G-150 Sephadex gel, resulting in an increase in the enzyme’s specific activity post-purification. It is well-understood that enzymes exhibit peak activity within specific, optimal conditions; deviations from these conditions can lead to diminished enzymatic performance.55
Application in Treatment of Diseases
Enzymes derived from fungi and bacteria, including cellulases, have therapeutic potential in managing conditions such as Phytobezoars, which result from the ingestion of indigestible plant matter causing blockages in the digestive tract. They also play a role in treating infections like blinding keratitis and granulomatous amoebic encephalitis, caused by pathogens such as Acanthamoeba, through the breakdown of plant cell walls and biofilms. Immobilized cellulases have applications beyond disease treatment; they function in bioreactors for enzyme deficiency disorders, serve as biosensors in bioremediation, assist in analytical flow systems, and contribute to the diagnosis and management of various medical conditions.
In the realm of commercial enzymatic preparations, cellulase from the natural fermentation of microorganisms like Trichoderma reesei and Bacillus licheniformis is a key component. These enzyme blends are engineered to facilitate the digestion of cellulose-rich dietary fibers found in fruits, vegetables, cereals, legumes, bran, nuts, seeds, soy, dairy products, greens, sprouts, and herbs. They also aid in the breakdown of fats, sugars, proteins, carbohydrates, and gluten.56
Fungal cellulases, when used in conjunction with enzymes like chitinases and lysozymes, have demonstrated efficacy in chitosan degradation. Chitosan, a valuable semicrystalline derivative of chitin, is obtained through partial deacetylation of chitin under alkaline conditions or enzymatic hydrolysis. Chitin, a structural polysaccharide present in the exoskeletons of marine organisms such as shrimp and in fungi, is transformed into chitosan, which has extensive medical applications including in surgical sutures, bone regeneration, production of artificial skin, and as an anticoagulant, antibacterial agent, and for cancer and diabetes management. Chitosan and its derivatives are also utilized in cosmetic formulations, biopharmaceutical production, and material encapsulation.57
Future Directions for Clinical Applications
Pythium being non-responsive to antifungals is a major hindrance to getting rid of the organism from the ocular surface. To circumvent the need for a therapeutic penetrating keratoplasty, other avenues of treatment should be looked for. Cellulose is an important structure providing stability to the plant cell wall. As mentioned earlier, cellulase has been used to break down cellulose and is utilized in various industries. Cellulase has a lytic action on the cell wall for protein and DNA extraction. Looking at its role in various sectors, we can remember that it could be useful in treating aggressive Pythium insidiosum keratitis. What can be kept in mind is the use of PHMB in Acanthamoeba keratitis. PHMB is a polymeric biguanide that is used as a disinfectant, especially in swimming pools. Larkin et al diluted PHMB and made it into a concentration of 0.02%. They showed complete resolution of infection in 5 patients with acanthamoeba keratitis in 1992. This led to the extensive use of PHMB in Acanthamoeba keratitis, which is a standard treatment at present. Although, still a large number of case still require keratoplasty due the severity of infection and virulence of the microorganism. Similarly, the use of cellulase in Pythium keratitis can be considered. It has to be subjected to quantitative and qualitative tests followed by toxicology and animal testing before it can be used in humans.
Acknowledgements
Sankara Nethralaya Medical Research Foundation, Chennai, Tamil Nadu, 600006, India.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis, and interpretation, or in all these areas; took part in drafting, revising, or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Gurnani B, Kaur K, Venugopal A., et al. Pythium insidiosum keratitis - A review. Indian J Ophthalmol. 2022;70(4):1107–1120. doi:10.4103/ijo.IJO_1534_21
2. Gurnani B, Christy J, Narayana S, Rajkumar P, Kaur K, Gubert J. Retrospective multifactorial analysis of Pythium keratitis and review of literature. Indian J Ophthalmol. 2021;69(5):1095–1101. doi:10.4103/ijo.IJO_1808_20
3. Gurnani B, Kaur K. Predicting prognosis based on regional prevalence, ulcer morphology and treatment strategy in vision-threatening Pythium insidiosum keratitis. Clin Ophthalmol. 2023;17:1307–1314. doi:10.2147/OPTH.S412274
4. Hasika R, Lalitha P, Radhakrishnan N, Rameshkumar G, Prajna NV, Srinivasan M. Pythium keratitis in South India: incidence, clinical profile, management, and treatment recommendation. Indian J Ophthalmol. 2019;67(1):42–47. doi:10.4103/ijo.IJO_445_18
5. Sahoo S, Mitra S, Mittal R, Behera HS, Das S. Use of different stains for microscopic evaluation for the diagnosis of Pythium keratitis. Indian J Med Microbiol. 2022;40(4):521–524. doi:10.1016/j.ijmmb.2022.08.003
6. Gurnani B, Kaur K, Agarwal S, et al. Pythium insidiosum Keratitis: past, Present, and Future. Ophthalmol Ther. 2022;11(5):1629–1653. doi:10.1007/s40123-022-00542-7
7. Bagga B, Sharma S, Madhuri Guda SJ, et al. Leap forward in the treatment of Pythium insidiosum keratitis. Br J Ophthalmol. 2018;102(12):1629–1633. doi:10.1136/bjophthalmol-2017-311360
8. Vishwakarma P, Mohanty A, Kaur A, et al. Pythium keratitis: clinical profile, laboratory diagnosis, treatment, and histopathology features post-treatment at a tertiary eye care center in Eastern India. Indian J Ophthalmol. 2021;69(6):1544–1552. doi:10.4103/ijo.IJO_2356_20
9. Gurnani B, Kaur K, Kumar T. Commentary: current concepts, recent updates, and future treatment options for Pythium insidiosum keratitis. Indian J Ophthalmol. 2023;71(5):1874–1876. doi:10.4103/IJO.IJO_80_23
10. Popper ZA, Ralet M-C, Domozych DS. Plant and algal cell walls: diversity and functionality. Ann Bot. 2014;114(6):1043–1048. doi:10.1093/aob/mcu214
11. Picard K, Tirilly Y, Benhamou N. Cytological effects of cellulases in the parasitism of phytophthora parasitica by pythium oligandrum. Appl Environ Microbiol. 2000;66(10):4305–4314. doi:10.1128/AEM.66.10.4305-4314.2000
12. García-Angulo P, Alonso-Simón A, Encina A, Álvarez JM, Acebes JL. Cellulose biosynthesis inhibitors: comparative effect on bean cell cultures. Int J Mol Sci. 2012;13(3):3685–3702. doi:10.3390/ijms13033685
13. Gaastra W, Lipman LJ, De Cock AW, et al. Pythium insidiosum: an overview. Vet Microbiol. 2010;146(1–2):1–16. doi:10.1016/j.vetmic.2010.07.019
14. Lekhanont K, Chuckpaiwong V, Chongtrakool P, Aroonroch R, Vongthongsri A. Pythium insidiosum keratitis in contact lens wear: a case report. Cornea. 2009;28(10):1173–1177. doi:10.1097/ICO.0b013e318199fa41
15. Reinprayoon U, Permpalung N, Kasetsuwan N, Plongla R, Mendoza L, Chindamporn A. Lagenidium sp. ocular infection mimicking ocular pythiosis. J Clin Microbiol. 2013;51(8):2778–2780. doi:10.1128/JCM.00783-13
16. Gurnani B, Kaur K. Pythium Keratitis. In: StatPearls. Treasure Island (FL): StatPearls Publishing; 2023.
17. Chatterjee S, Agrawal D, Gomase SN. Clinical differentiation of Pythium keratitis from fungal keratitis and development of a scoring system. Indian J Ophthalmol. 2022;70(10):3515–3521. doi:10.4103/ijo.IJO_870_22
18. Kalra P, Bagga B, Garg P. Pythium insidiosum keratitis: histopathology and rapid novel diagnostic staining technique. Cornea. 2018;37(3):e14. doi:10.1097/ICO.0000000000001463
19. Agarwal S, Iyer G, Srinivasan B, et al. Clinical profile, risk factors and outcome of medical, surgical and adjunct interventions in patients with Pythium insidiosum keratitis. Br J Ophthalmol. 2019;103(3):296–300. doi:10.1136/bjophthalmol-2017-311804
20. Cosgrove DJ. Building an extensible cell wall. Plant Physiol. 2022;189(3):1246–1277. doi:10.1093/plphys/kiac184
21. Tateno M, Brabham C, DeBolt S. Cellulose biosynthesis inhibitors - A multifunctional toolbox. J Exp Bot. 2016;67(2):533–542. doi:10.1093/jxb/erv489
22. Latgé JP. The cell wall: a carbohydrate armour for the fungal cell. Mol Microbiol. 2007;66(2):279–290. doi:10.1111/j.1365-2958.2007.05872.x
23. Blaschek W, Käsbauer J, Kraus J, Franz G. Pythium aphanidermatum: culture, cell-wall composition, and isolation and structure of antitumour storage and solubilised cell-wall (1----3),(1----6)-beta-D-glucans. Carbohydr Res. 1992;231:293–307. doi:10.1016/0008-6215(92)84026-o
24. Yamada M, Miyazaki T. Ultrastructure and chemical analysis of the cell wall of Pythium debaryanum. Jpn J Microbiol. 1976;20(2):83–91. doi:10.1111/j.1348-0421.1976.tb00913.x
25. Natarajan R, Janani MK. Tissue Adhesive to Tackle a Terrible Infection in Vitro Inhibition of Pythium Insidiosum Keratitis Isolates By 1 N-Butyl-2-Cyanoacrylate. Ophthalmic Pathol. 2021;3(3):123.
26. Kraehmer H, Laber B, Rosinger C, Schulz A. Herbicides as weed control agents: state of the art: i. Weed control research and safener technology: the path to modern agriculture. Plant Physiol. 2014;166(3):1119–1131. doi:10.1104/pp.114.241901
27. Grossmann K, Tresch S, Plath P. Triaziflam and Diaminotriazine derivatives affect enantioselectively multiple herbicide target sites. Z Naturforsch C J Biosci. 2001;56(7–8):559–569. doi:10.1515/znc-2001-7-814
28. Brabham C, Lei L, Gu Y, Stork J, Barrett M, DeBolt S. Indaziflam herbicidal action: a potent cellulose biosynthesis inhibitor. Plant Physiol. 2014;166(3):1177–1185. doi:10.1104/pp.114.241950
29. Gu Y, Kaplinsky N, Bringmann M, et al. Identification of a cellulose synthase-associated protein required for cellulose biosynthesis. Proc Natl Acad Sci U S A. 2010;107(29):12866–12871. doi:10.1073/pnas.1007092107
30. Drakakaki G, van de Ven W, Pan S, et al. Isolation and proteomic analysis of the SYP61 compartment reveal its role in exocytic trafficking in Arabidopsis [published correction appears in Cell Res. 2023 Sep 21;:]. Cell Res. 2012;22(2):413–424. doi:10.1038/cr.2011.129
31. Xia Y, Lei L, Brabham C, et al. Acetobixan, an inhibitor of cellulose synthesis identified by microbial bioprospecting. PLoS One. 2014;9(4):e95245. doi:10.1371/journal.pone.0095245
32. Deguchi S, Degaki H, Taniguchi I, Koga T. Deep-sea-inspired chemistry: a hitchhiker’s guide to the bottom of the ocean for chemists. Langmuir. 2023;39(23):7987–7994. doi:10.1021/acs.langmuir.3c00516
33. Kumar R, Singh S, Singh OV. Bioconversion of lignocellulosic biomass: biochemical and molecular perspectives. J Ind Microbiol Biotechnol. 2008;35(5):377–391. doi:10.1007/s10295-008-0327-8
34. Dhiman TR, Zaman MS, Gimenez RR, Walters JL, Treacher R. Performance of dairy cows fed forage treated with fibrolytic enzymes prior to feeding. Anim Feed Sci Technol. 2002;101(1):115–125.
35. Godfrey T, West S. “Textiles”, in Industrial Enzymology.
36. Bhat MK. Cellulases and related enzymes in biotechnology. Biotechnol Adv. 2000;18(5):355–383.
37. Sukumaran RK, Singhania RR, Pandey A. Microbial cellulases—production, applications and challenges. J Sci Ind Res. 2005;64(11):832–844.
38. Hebeish A, Ibrahim NA. The impact of frontier sciences on textile industry. Colourage. 2007;54:41–55.
39. Galante YM, DeConti A, Monteverdi R. “Application of Trichoderma enzymes in food and feed industries”, in Tri- choderma and Gliocladium—Enzymes. In: Harman GF, Kubicek CP, editors. Biological Control and Commercial Applications. Vol. 2. London, UK: Taylor & Francis; 1998:311–326.
40. Balouiri M, Sadiki M, Ibnsouda SK. Methods for in vitro evaluating antimicrobial activity: a review. J Pharm Anal. 2016;6(2):71–79.
41. Dogan C, Aygun G, Bahar-Tokman H, et al. In vitro antifungal effect of acrylic corneal glue (N-Butyl-2-Cyanoacrylate). Cornea. 2019;38(12):1563–1567.
42. Petrikkou E, Rodríguez-Tudela JL, Cuenca-Estrella M, Gómez A, Molleja A, Mellado E. Inoculum standardization for antifungal susceptibility testing of filamentous fungi pathogenic for humans. J Clin Microbiol. 2001;39(4):1345–1347. doi:10.1128/JCM.39.4.1345-1347.2001
43. Romero IL, Malta JB, Silva CB, Mimica LM, Soong KH, Hida RY. Antibacterial properties of cyanoacrylate tissue adhesive: does the polymerization reaction play a role? Indian J Ophthalmol. 2009;57(5):341–344. doi:10.4103/0301-4738.55065
44. Henrissat B, Teeri TT, Warren RAJ. A scheme for designating enzymes that hydrolyse the polysaccharides in the cell walls of plants. FEBS Lett. 1998;425(2):352–354.
45. Sang-Mok L, Koo YM. Pilot-scale production of cellulase using Trichoderma reesei Rut C-30 in fed-batch mode. J Microbiol Biotechnol. 2001;11(2):229–233.
46. Kuhad RC, Manchanda M, Singh A. Hydrolytic potential of extracellular enzymes from a mutant strain of Fusarium oxysporum. Bioprocess Eng. 1999;20(2):133–135.
47. Herbert R, Sharp R. Chapman and Hall. Molecular Biology and Biotechnology of Extremophiles; 1992.
48. Kubicek CP, Penttila ME. Regulation of production of plant polysaccharide degrading enzymes by Trichoderma. In: Harman E, Kubicek CP, editors. In Trichoderma and Gliocladium. London: Taylor & Francis Ltd; 1998:49–72.
49. Aro N, Ilmen M, Saloheimo A, Penttila M. ACEI is a repressor of cellulase and xylanase genes of Trichoderma reesei. Appl Environ Microbiol. 2002;69:56–65.
50. Strauss J, Mach RL, Zeilinger S, et al. Cre1, the carbon catabolites repressor protein from Trichoderma reesei. FEBS Letts. 1995;376:103–107.
51. Narendja FM, Davis MA, Hynes MJ. An CF, the CCAAT binding complex of Aspergillus nidulans, is essential for the formation of a DNAse I-hypersensitive site in the 50 region of the amdS gene. Mol Cell Biol. 1999;19:6523–6531.
52. Suto M, Tomita F. Induction and catabolic repression mechanisms of cellulase in fungi. J Biosci Bioeng. 2001;92:305–311.
53. Inoue T, Moriya S, Ohkuma M, Kudo T. Molecular cloning and characterization of a cellulase gene from a symbiotic protist of the lower termite, Coptotermes formosanus. Gene. 2005;349:67–75.
54. Silano V, Barat Baviera JM, Bolognesi C, et al. EFSA Panel on Food Contact Materials, Enzymes and Processing Aids (CEP). Safety evaluation of the food enzyme cellulase from Trichoderma reesei (strain DP-Nzc36). EFSA J. 2019;17:10.
55. Worden N, Wilkop TE, Esteve VE, et al. CESA TRAFFICKING INHIBITOR inhibits cellulose deposition and interferes with the trafficking of cellulose synthase complexes and their associated proteins KORRIGAN1 and POM2/CELLULOSE SYNTHASE INTERACTIVE PROTEIN1. Plant Physiol. 2015;167(2):381–393. doi:10.1104/pp.114.249003
56. Aro N, Saloheimo A, Ilmen M, Penttila M. ACEII, a novel transcriptional activator involved in regulation of cellulase and xylanase genes of Trichoderma reesei. J Biol Chem. 2001;276:24309–24314.
57. Xin X, Lei L, Zheng Y, et al. Cellulose synthase interactive1- and microtubule-dependent cell wall architecture is required for acid growth in Arabidopsis hypocotyls. J Exp Bot. 2020;71(10):2982–2994. doi:10.1093/jxb/eraa063
 © 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Predicting Prognosis Based on Regional Prevalence, Ulcer Morphology and Treatment Strategy in Vision-Threatening Pythium insidiosum Keratitis
Gurnani B, Kaur K
Clinical Ophthalmology 2023, 17:1307-1314
Published Date: 5 May 2023