")
Back to Journals » International Journal of Chronic Obstructive Pulmonary Disease » Volume 18
MEG3 Regulates CSE-Induced Apoptosis by Regulating miR-421/DFFB Signal Axis
Authors Bi H, Wang G, Li Z, Zhou L, Zhang M
Received 20 January 2023
Accepted for publication 30 April 2023
Published 15 May 2023 Volume 2023:18 Pages 859—870
DOI https://doi.org/10.2147/COPD.S405566
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 6
Editor who approved publication: Dr Richard Russell
Hui Bi,1 Gui Wang,2 Zhiying Li,1 Lin Zhou,1 Ming Zhang1
1Department of Respiratory Medicine, The Third Affiliated Hospital of Soochow University, Changzhou, Jiangsu, People’s Republic of China; 2Department of Intensive Care Unit, The Third Affiliated Hospital of Soochow University, Changzhou, Jiangsu, People’s Republic of China
Correspondence: Hui Bi, Department of Respiratory Medicine, The Third Affiliated Hospital of Soochow University, Changzhou, Jiangsu, 213000, People’s Republic of China, Email [email protected]
Introduction: Chronic obstructive pulmonary disease (COPD) is a common respiratory disease with irreversible and progressive obstruction of airflow. Currently, there are no clinically available treatments to prevent COPD progression. Apoptosis of human lung microvascular endothelial cells (HPMECs) and bronchial epithelial cells (HBECs) is often observed in COPD, but its pathogenesis has not been fully elucidated. LncRNA maternally expressed gene 3 (MEG3) is closely related to CSE-induced apoptosis, but the specific mechanism of MEG3 in COPD is still unknown.
Methods: In the present study, cigarette smoke extract (CSE) is used to treat HPMECs and HBECs. Flow cytometry assay is used to detect the apoptosis of these cells. The expression of MEG3 in CSE-treated HPMECs and HBECs is detected by qRT-PCR. LncBase v.2 is used to predict miRNAs binding to MEG3, and miR-421 is found to bind to MEG3. Dual luciferase report analysis and RNA immunoprecipitation experiment jointly clarified the binding relationship between MEG3 and miR-421.
Results: MiR-421 was downregulated in CSE-treated HPMECs/HBECs, and miR-421 overexpression mitigated CSE-induced apoptosis in these cells. Subsequently, DFFB was found to be directly targeted by miR-421. The overexpression of miR-421 dramatically reduced the expression level of DNA fragmentation factor subunit beta (DFFB). DFFB was found downregulated in CSE-treated HPMECs and HBECs. MEG3 contributed to the apoptosis of HPMECs and HBECs induced by CSE by regulating the miR-421/DFFB axis.
Conclusion: This study presents a new perspective on the diagnosis and treatment of COPD caused by CSE.
Keywords: COPD, CSE, MEG3, miR-421/DFFB axis, apoptosis
Introduction
Among chronic respiratory diseases, COPD is very common and has characteristics of irreversible lung failure with high morbidity and mortality worldwide, causing a huge economic and social burden.1 With the increasing incidence, COPD is becoming the third most fatal disease on the planet.2 The epidemiological survey of COPD in China published in the Lancet in 2018 found that the prevalence rate of COPD among people aged 40 and above was 13.7%. WHO predicts that by 2030, 4.5 million people will die from COPD each year, making it a major global public health problem.3 Previous studies have shown that the pathogenesis of COPD is related to a variety of factors, such as environmental risk,4 abnormal inflammatory response,5 and genetic susceptibility.6
Research indicated that cigarette smoke (CSE) is one of the important factors that promote the occurrence and development of COPD. CSE can induce apoptosis, endothelial damage, and oxidative stress, and ultimately lead to continuous inflammation of lung tissue.7 Glucocorticoids and bronchodilators are currently the main treatments for COPD, which can improve symptoms, but the effect is limited. No treatment has been demonstrated to reduce the progression of the disease.8 Therefore, it is critical to investigate its possible molecular process.
LncRNAs are noncoding RNAs that cannot encode proteins (200 nt in length). They can serve as competitive endogenous (CE) RNA and play an important role in regulating gene expression through sponge miRNAs in various physiological and pathological processes.9 A previous study elucidates that lncRNA MIR155HG /miR-128-5p participates in smoking-induced COPD by regulating the expression of BRD4.10 LncRNA NNT-AS1 regulates the apoptosis and inflammation of COPD by regulating the miR-582-5p/FBXO11 axis.11 LINC00612 participates in CSE-induced apoptosis of HPMECs by regulating apoptosis, inflammation, and oxidative stress.12 In cigarette smoke-induced COPD, lncRNA taurine upregulated gene 1 (TUG1) promotes airway remodeling.13 Previous research has clarified that MEG3 is involved in regulating the apoptosis of human pulmonary microvascular endothelial cells (HPMECs) in COPD.6 In a COPD cell model constructed from human bronchial epithelial cells (16HBE) induced by CSE, the expression level of MEG3 is significantly upregulated, and knockdown of MEG3 can promote cell proliferation through negative regulation of miR-181a-2-3p and miR-218, and inhibit cell apoptosis and inflammation levels.14,15 These studies preliminarily clarified the important role of MEG3 in COPD. However, the exact regulation mechanism of MEG3 in the occurrence and development of COPD needs to be further explored.
MiRNAs are short non-coding RNAs (length 19–25 nt) and participate in the regulation of diseases by regulating the expression of protein-coding genes.16 So far, more and more reports have clarified that miRNAs play a key role in the occurrence and development of COPD. For example, miR-150 negatively regulates the expression of P53 to inhibit CS-induced lung inflammation and airway epithelial cell apoptosis.17 MiR-145-5p partially protects CSE-induced apoptosis of airway epithelial cells by targeting KLF5.18 An et al found that miR-421 may play a role in COPD through network analysis.19 However, the molecular mechanism of miR-421 in COPD has not been reported yet.
DNA fragmentation factor subunit beta (DFFB) is the main intracellular nuclease involved in DNA fragmentation in the process of cell apoptosis, and plays an important role in DNA fragmentation in the process of cell apoptosis.20 Research has shown that tobacco compounds can affect apoptosis in human fetal lung cell line (WI38) by regulating the expression of DFFB.21 However, it has not been reported whether DFFB is involved in regulating cell apoptosis in COPD.
HPMECs and human bronchial epithelial cells (HBECs) are commonly used cell types in COPD research.6,14 HPMECs is a kind of endothelial cells present in human lung tissue, which have the roles of maintaining the normal function of alveoli, secreting mucus. HBECs is a type of cell present on the bronchial mucosal surface, which plays a role in maintaining the normal function of the trachea and secreting mucus.22,23 HMEC and HBE have important roles in the study of COPD, and through the study of these two types of cells, we can help us better understand the pathogenesis and treatment of COPD. In this study, we aimed to investigate the molecular mechanism of the ceRNA regulatory network of MEG3/miR-421/DFFB in CSE-stimulated HPMECs and HBECs.
Materials and Methods
Cell Culture Conditions and CSE-Treatment
Human pulmonary microvascular endothelial cells (HPMECs) and human bronchial epithelial (HBECs) were purchased from China Cell Research Institute Center (Shanghai). HPMECs were grown in endothelial cell medium (ECM, Promocell) (adding 1% endothelial cell growth factor, 5% FBS, 100 U/mL penicillin, and 50 μg/mL streptomycin). HBECss were cultured in DMEM/F-12 (Wisent, Canada) (adding 1% endothelial cell growth factor, 5% FBS, 100 U/mL penicillin, and 50 μg/mL streptomycin). All these cells were cultured with 5% CO2 at 37 °C. Besides, according to previous reports, CSE was prepared from cigarette extracts.14,24 Shortly, smoke from ten cigarettes (Hunan Zhongyan Industrial Co., Ltd.) is dissolved in 25 mL of phosphate buffered saline (PBS). The suspension was adjusted to pH 7.2–7.4 and filtered using a cellulose membrane (0.22 µm) to remove bacteria. The solution was treated as 100% CSE and diluted with PBS to obtain a concentration of 5%. To study COPD in vitro, in CSE group, we treated HPMECs and HBECs cells with 5% CSE for 24 h. In the control group, the HPMECs and HBECs cells was treated with the PBS.
Plasmid Construction, Lentivirus Production, and Cell Transduction
The pcDNA3.1 of MEG3 (including pcDNA3.1 vector), the siRNA of MEG3 (including negative control (NC)-siRNA), and The lentiviral vector expressing short hairpin RNA (siRNA) (including NC-siRNA) targeting MEG3 or DFFB was designed and constructed by GenePharma (Shanghai, China) and transfected into HPMECs and HBECs with Lipofectamine 2000 reagents (Invitrogen). In addition, the miR-421 mimics/inhibitors (including NC-mimics/inhibitors) were also purchased from GenePharma (Shanghai, China). After 48 h transfection, the HPMECs and HBECs were used in subsequent experiments.
Cell Apoptosis
After transfection, HPMECs and HBECs were inoculated in cell culture plates for 12 h. After the number of cells reached 5×105, these cells were collected and washed with pre-cooled PBS. Then add 5 μL Annexin V-FITC and 5 μL propidium iodide (PI) staining, mixing, avoiding light, incubating at room temperature for 15 minutes. Collect data on FACSVerse flow cytometry (BD Biosciences) and analyze using FlowJo software (TreeStar, Inc).
RNA Extraction and qRT-PCR
The qRT-PCR experiment is described in previous reports.25 In brief, the total RNA of HPMECs/HBECs was isolated by TRIzol reagent (Invitrogen). The PrimeScipt RT Master Mix kit (Takara, Tokyo, Japan) was used to prepare cDNA. The SYBR® Premix Ex TaqTM reagent (TaKaRa) was used to quantitatively analyze gene expression. The 2−∆∆Ct method was used to calculate the relative expression of MEG3/miR-421/DFFB. GAPDH and U6 snRNA served as the normalized controls. The primers for MEG3, miR-421, and DFFB were purchased from GenePharma, and the specific sequences were as followed: U6: F: 5’- CTCGCTTCGGCAGCACA-3’, R: 5’- AACGCTTCACGAATTTGCGT-3’. MEG3: F: 5’-CAGGATGGCAAAGGATGAAG-3’, R: 5’-GCAGGTGAACACAAGCAAAGA-3’. miR-421: F: 5’- GTCGCGCGGGUUAAUGCCTC-3’, R: 5’- GGACATUAGUUGUCUGUAAATAG-3’. DFFB: F: 5’- CACAACGTCAGCCAGAACAT-3’, R: 5’- CCCAGTCCACTTCTCTTCCA-3’. GAPDH: F: 5’-TGTTCGTCATGGGTGTGAAC-3’, R: 5’- ATGGCATGGACTGTGGTCAT-3’.
Luciferase Reporter Gene Detection
The Luciferase reporter plasmid of pMIR-MEG3-WT/pMIR-MEG3-MUT and the Luciferase reporter plasmid of pMIR-DFFB-WT/pMIR-DFFB-MUT were constructed, and the plasmid was transfected into HPMECs and HBECs with miR-421/NC mimic. After 48 h, the Luciferase signal was detected by the Luciferase detection kit, and the differences in Luciferase signal between different groups were compared.
RNA Immunoprecipitation Assay
The Biotin-labeled MEG3 probe was constructed according to the kit instructions. The cells induced by hyperoxia were collected, fixed with formaldehyde, lysed with RIP lysate, and sonicated, centrifuged at 1000 g for 20 min, and the supernatant was obtained as Input. The precipitate was incubated with magnetic beads coupled with a Biotin-labeled MEG3 probe overnight, centrifuged to wash, and the supernatant was removed to obtain a protein-RNA complex. After that, it was incubated with proteinase K at 55 °C for 30 min, uncoupling was performed, and RNA was collected and detected by qRT-PCR.
Western Blotting
Collect cells and lyse them in RIPA lysis buffer (Beyotime, Shanghai, China). Then, the total protein was quantified using the BCA protein assay kit (Thermo Scientific). The same amount of protein was separated on SDS polyacrylamide gel and transferred to polyvinylidene fluoride membrane (PVDF) (Millipore, Billerica, USA). Seal with 5% skimmed milk at room temperature for 1 hour. Wash with TBST, and use the primary antibody anti-β-Actin (1:2000), anti-DNA fragmentation factor subunit beta (DFFB) (1:1000), anti-argonaute-2 (Ago2) (1:1000) incubate overnight at 4 °C. Wash the membrane with TBST 3 times (15 min/time), then incubated with secondary antibody diluent (1:2000) for 2 h, then wash 3 times with TBST (15 min/time), and finally develop color with Tanon Luminescence Kit. Record the imaging results.
Statistical Analyses
All experiments are set up for at least 3 biological replicates and then averaged. The mean value of each experiment is expressed as mean±standard deviation (SD). The Student’s t-test was used to compare two samples. Statistical differences between three or more groups of data were evaluated using the one-way ANOVA test, followed by the Tukey post-test. P value <0.05 was considered significant.
Results
MEG3 is Significantly Upregulated During CSE-Induced Apoptosis of HPMECs and HBECs Cells
To study the mechanism of MEG3 in CSE-induced COPD, we first used 5% CSE to process HPMECs and HBECs to construct a COPD cell model. Then, flow cytometry was used to detect the apoptosis ratio of HPMECs and HBECs treated with CSE. As shown in Figure 1A, compared with the control group, the apoptosis rate of HPMECs and HBECs treated with CSE was significantly increased. At last, qRT-PCR was used to detect the expression level of MEG3 in HPMECs and HBECs. The data showed that the expression of MEG3 in HPMECs and HBECs cells treated with CSE was significantly upregulated compared with the control group (Figure 1B). These results suggest that MEG3 may be involved in regulating the process of apoptosis induced by CSE in HPMECs and HBECs.
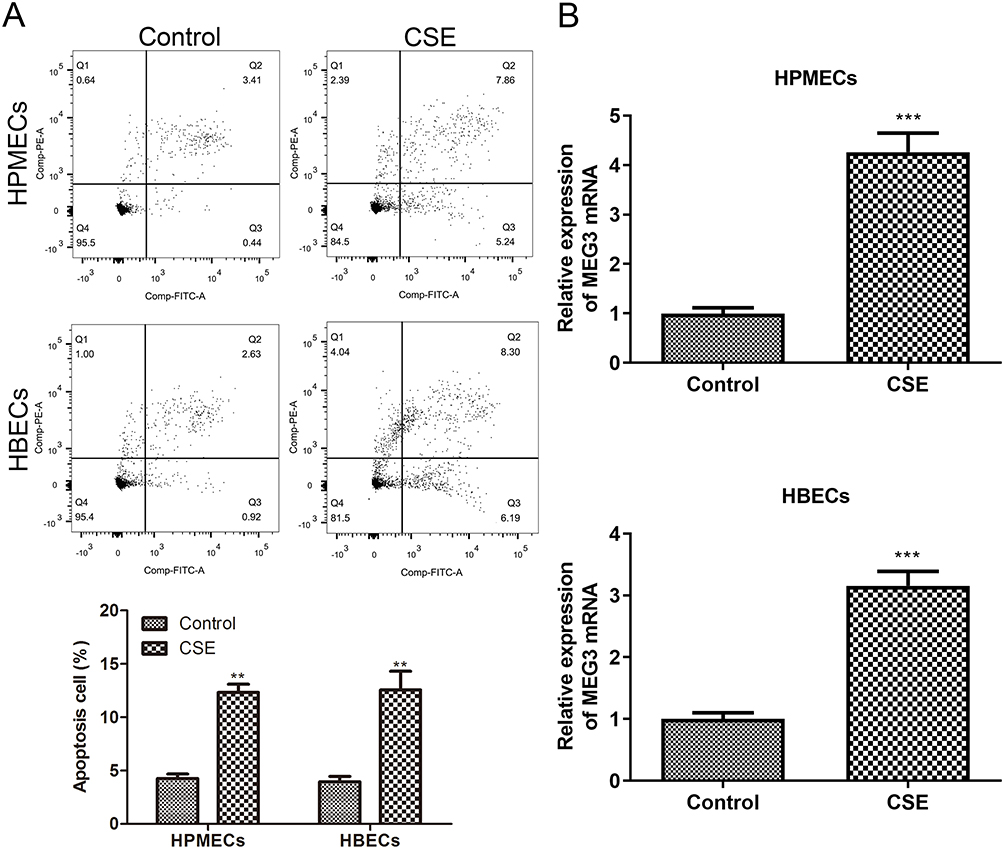 |
Figure 1 The expression of MEG3 during CSE-induced apoptosis of HPMECs and HBECs. (A) 5% CSE processes HPMECs and HBECs, and uses flow cytometry to detect the rate of apoptosis. (B) RT-qPCR was used to detect MEG3 expression in HPMECs and HBEC with or without 5% CSE treatment. The experiment was repeated three times and then averaged. (**p < 0.01; ***p < 0.001). |
MEG3 Promotes the Apoptosis of HPMECS and HBECs Treated with CSE
To further clarify the effect of MEG3 on the apoptosis of HPMECs and HBECs, we first constructed a MEG3 overexpression plasmid using pcDNA3.1 vector and then transfected it into HPMECs and HBECs to overexpress MEG3. Then, qRT-PCR was used to detect overexpression efficiency. Compared with the control group, the expression of MEG3 in the MEG3 overexpression group (pc-MEG3) was significantly upregulated (Figure 2A). Next, we tested the apoptosis rate of these cells. The data showed that overexpression of MEG3 significantly increased the apoptotic ratio of HPMECs and HBECs (Figure 2B).
 |
Figure 2 The overexpression of lncRNA MEG3 affects the apoptosis of HPMEC and HBEC. (A) After transfection with pcDNA3.1 vector or pcDNA3.1 MEG3, the mRNA expression level of MEG3 in HPMECs and HBEC was determined by qRT-PCR. (B) The apoptosis rates of HPMECs and HBECs transfected with pcDNA3.1 vector or pcDNA3.1 MEG3 were analyzed by flow cytometry. The experiment was repeated three times and then averaged. (***p < 0.001). |
In addition, we synthesized MEG3 siRNA and transfected them into HPMECs and HBECs treated with CSE to knock down the expression of MEG3. As shown in Figure 3A, knocking down of MEG3 significantly reduced the expression of MGE3. Finally, we tested the apoptosis of these cells treated with CSE. The results were consistent with expectations. Knockdown of MEG3 significantly reduced the apoptotic ratio of HPMECs and HBECs treated with CSE (Figure 3B). All these results indicated that MEG3 advanced the apoptosis of HPMECS and HBECs treated with CSE.
 |
Figure 3 The effect of lncRNA-MEG3 knockdown on the apoptosis of HPMECs and HBECs. (A) After transfection with NC-siRNA or MEG3-siRNA, the expression level of MEG3 mRNA in HPMECs and HBECs treated with CSE was determined by qRT-PCR. (B) Apoptosis of HPMEC and HBECs treated with CSE after transfection with NC-siRNA or MEG3-siRNA was determined by flow cytometry. The experiment was repeated three times and then averaged. (**p < 0.01; ***p < 0.001). |
MiR-421 is a Target Gene of MEG3 in HPMECS and HBECs
A multitude of studies showed that lncRNAs participated in the regulation of various physiological and pathological processes through sponging miRNA. Thus, LncBase v.2 was used to predict the target miRNAs of MGE3. As shown in Figure 4A, miR-421 and MEG3 had binding regions. Then, the dual luciferase reporter assay was used to validate the potential interaction between miR-421 and MEG3. We constructed MEG3-WT or MEG3-MUT luciferase reporter plasmids and transfected these plasmids into HPMECS and HBECs together with NC mimic and miR-421 mimic. The results showed that MEG3 targeted miR-421 (Figure 4B). In addition, we further clarified the interaction of MEG3 and miR-421 in the same RNA-induced silencing complex (RISC) through RIP assay. Western blot was used to detect the RIP efficiency of Ago2 protein (Figure 4C). MEG3 and miR-421 levels in the anti-ago2 group were significantly increased compared with the anti-IgG group (Figure 4D). Finally, we tested the expression changes of miR-421 after CSE treated these cells. The results showed that CSE-treated cells significantly reduced the expression of miR-421 (Figure 4E). Subsequently, we further used MEG3 siRNA to stably knockdown the expression of MEG3 in HPMECs and HBECs treated with CSE, and detected the expression of miR-421. The results showed that knocking down MEG3 significantly increased the expression of miR-421 (Figure 4F). All in all, these results indicated that MEG3 is directly bound to miR-421 in HPMECs and HBECs.
 |
Figure 4 MiR-421 targets MEG3 in HPMECs and HBECs. (A) Diagram of MEG3 and miR-421 binding sites. (B) In HPMEC and HBEC, the effect of miR-421 overexpression on MEG3-WT/MEG3-MUT reporter luciferase activity was evaluated by dual luciferase reporter gene analysis. (C) Western blot was used to detect the RIP efficiency of Ago2 protein. (D) The cell lysate was used to perform the RIP experiments. The qRT-PCR was used to detect the MEG3 and miR-421 expression in HPMECs and HBECs. (E) miR-421 expression in HPMECs and HBECs was detected by qRT-PCR when treated with or without 5% CSE. The experiment was repeated three times and then averaged. (F) After transfection with si-NC or si-MEG3, the expression level of miR-421 in HPMEC and HBEC treated with CSE was detected by qRT-PCR. (**p < 0.01; ***p < 0.001). |
Overexpression of miR-421 Significantly Reduces CSE-Induced Apoptosis in HPMECs and HBECs Cells
To further explore the role of miR-421 in the apoptosis of HPMECs and HBECs induced by CSE, we first used miR-421 mimic to transfect these cells to overexpress miR-421, and tested the overexpression efficiency of miR-421 (Figure 5A). Then, we tested the apoptotic ratio of HPMECs and HBECs treated with CSE after miR-421 was overexpressed. The results showed that overexpression of miR-421 significantly reduced CSE-induced apoptosis (Figure 5B). These results indicated that miR-421 negatively regulated CSE-induced apoptosis in HPMECs and HBECs.
 |
Figure 5 The effect of miR-421 overexpression on the apoptosis of HPMECs and HBECs. (A) The expression of miR-421 in HPMECs and HBECs treated with CSE was detected by qRT-PCR after transfection with either NC mimic or miR-421 mimic. (B) 5% CSE processes HPMECs and HBECs after transfection with NC mimic or miR-421 mimic, and uses flow cytometry to detect the rate of apoptosis. The experiment was repeated three times and then averaged. (**p < 0.01; ***p < 0.001). |
MiR-421 Targets the 3’-UTR of DFFB
MiR-421 regulated the expression of target genes by regulating the 3’-UTR region of target genes and ultimately participated in the regulation of physiological and pathological processes. First, we screened miR-421 for a total of 50 downstream targets in ENCORI, miRWalk, and miRDB databases, and then selected DFFB by literature mining (Figure 6A). As shown in Figure 6B, miR-421 and DFFB have binding regions. The relationship between miR-421 and DFFB was subsequently verified through a luciferase reporter gene assay. We constructed luciferase reporter plasmids containing the normal or mutant DFFB 3’-UTR and transfected these plasmids into HPMECs and HBECs together with NC mimic and miR-421 mimic. The results indicated that miR-421 targeted the 3’-UTR of DFFB (Figure 6C). Besides, we further assessed DFFB protein expression in these cells. The results showed that DFFB protein expression was markedly decreased in the miR-421 overexpression group (Figure 6D). Finally, we detected the mRNA of DFFB in HPMECs and HBECs treated with CSE. The results showed that CSE significantly increased the expression of DFFB (Figure 6E). Subsequently, we tested the mRNA expression changes of DFFB after overexpression of miR-421. The results showed that overexpression of miR-421 would significantly reduce the mRNA expression of DFFB (Figure 6F). The above results indicated that miR-421 targeted and negatively regulated DFFB expression.
 |
Figure 6 MiR-421 targets DFFB expression. (A) The Venn diagram shows the miR-421 targets from the ENCORI, miRWalk, and miRDB databases. (B) The 3’-UTR base pairing diagram of miR-421 and DFFB. (C) In HPMEC and HBEC, the effect of miR-421 overexpression on DFFB-WT/DFFB-MUT reporter luciferase activity was evaluated by dual luciferase reporter gene analysis. (D) The DFFB protein expression in HPMECs and HBECs when transfection with NC mimic or miR-421 mimic was detected by WB. (E) The mRNA expression levels of DFFB in HPMECs and HBECs treated with or without 5% CSE was detected by qRT-PCR. (F) The mRNA expression levels of DFFB in HPMECs and HBECs treated with CSE were detected by qRT-PCR after transfection with NC mimic or miR-421 mimic. The experiment was repeated three times and then averaged. (**p < 0.01, ***p < 0.001 versus the NC mimic or Control). |
MEG3 Promoted CSE-Induced Apoptosis of HPMECs and HBECs by Regulating the Expression of DFFB Through Sponging miR-421
To further clarify the role of MEG3 in CSE-induced COPD, we first used a lentiviral vector expressing short hairpin RNA (shRNA) to construct the stable knockdown cell lines of MEG3 (si-MEG3) in HPMECs and HBECs. Then, we transfected NC-inhibitor or miR-421 inhibitor into these si-MEG3 cell lines. Subsequently, qRT-PCR was used to detect the expression of miR-421. As shown in Figure 7A, compared with the si-MEG3 cell lines transfected with NC-inhibitor, miR-421 were all significantly down-regulated in the si-MEG3 cell lines transfected with miR-421 inhibitor. In addition, we further detected the expression of DFFB in HPMECs and HBECs that both knocked down miR-421 and DFFB in si-MEG3 cell lines. The results showed that, compared with si-MEG3 cell lines transfected with NC-inhibitor+si-NC, DFFB were all significantly upregulated in the si-MEG3 cell lines transfected with miR-421 inhibitor+si-NC, while DFFB were all significantly down-regulated in the si-MEG3 cell lines transfected with miR-421 inhibitor+si-DFFB (Figure 7A). Finally, we examined the changes of apoptosis in these cells. The results showed that compared with si-MEG3 cell lines transfected with NC-inhibitor+si-NC, the apoptosis ratio was all significantly upregulated in the si-MEG3 cell lines transfected with miR-421 inhibitor+si-NC, while the apoptosis ratio was all significantly down-regulated in the si-MEG3 cell lines transfected with miR-421 inhibitor+si-DFFB (Figure 7B). These results indicated that MEG3 participated in the regulation of CSE-induced apoptosis of HPMECs and HBECs by regulating the miR-421/DFFB axis.
 |
Figure 7 The rescue experiment verified the function of MEG3/miR-421/DFFB in the CSE-induced apoptosis model. (A) The miR-421 expression of HPMECs and HBECs when transfection with sh-MEG3+NC inhibitor or sh-MEG3+miR-421 inhibitor. The DFFB mRNA expression of HPMECs and HBECs were detected by qRT-PCR after transfection with sh-MEG3+NC inhibitor+si-NC, sh-MEG3+miR-421 inhibitor+si-NC, and sh-MEG3+miR-421 inhibitor+si-DFFB. (B) After being treated with CSE, the cells were transfected with sh-MEG3+NC inhibitor+si-NC, sh-MEG3+miR-421 inhibitor+si-NC or sh-MEG3+miR-421 inhibitor+si-DFFB, the apoptosis of HPMEC and HBECs was detected by flow cytometry. The experiment was repeated three times and then averaged. (**p < 0.01, ***p < 0.001 versus the NC inhibitor or NC inhibitor+si-NC group; #p < 0.05, ##p < 0.01 versus the miR-421 inhibitor+si-NC group). |
Discussion
COPD is a multi-factor complex disease determined by genetic and environmental factors.26 Previous studies have shown that a very important predisposing factor in the progression of chronic obstructive pulmonary disease is smoking, and smoking regulates the apoptosis of HPMECs and HBECs.6,27 In addition, previous studies have found that increased apoptosis has been observed in COPD animals and patients.28,29 The results in this study are in accord with recent studies: the apoptosis rate of HPMECs and HBECs treated with 5% CSE was significantly increased.
LncRNAs have been reported to play important functions in many diseases, and the interaction of lncRNA-miRNA-mRNA may play key roles in COPD.12 LncRNA MEG3 is not only a regulator of cell biological behavior,30 but also a masked target of many diseases.31,32 Previous research has linked the lncRNA MEG3 to being down-regulated in melanoma, hepatocellular carcinoma, and Parkinson's disease.33–35 One unexpected finding was the extent to which, in this study, we found that MEG3 was significantly overexpressed in HPMECs and HBECs treated with CSE. In addition, the upregulation of MEG3 leads to more apoptosis of HPMECs and HBEC. However, si-MEG3 leads to decreased apoptosis of HPMECs and HBEC. Experimental data suggest that MEG3 may play a key role in COPD by regulating HPMECs and HBEC apoptosis.
Several studies have shown that lncRNAs exert their function by interacting with miRNAs.36,37 For example, LncRNA SNHG5 regulates the expression of PTEN through sponging miR-132 to regulate cell apoptosis in COPD.38 Besides, MEG3 is involved in a variety of diseases by serving as a sponge for miRNA. For instance, MEG3 participates in the regulation of CSE-induced apoptosis and inflammation through sponging miR‑181a‑2‑3p or miR-218 in 16HBE cells.14,15 In this study, we found that MEG3 targeted negative regulation of miR-421, which was significantly downregulated in CSE treated cells. Overexpression of miR-421 significantly reduced apoptosis induced by CSE.
Because miRNAs exert their biological functions by regulating their downstream targets.39 Previous studies have shown that the expression of miR-132 is upregulated in THP-1 and BEAS-2B cells treated with CSE, and increases the level of cytokines by targeting down regulation of SOCS5 expression.40 However, whether miR-421 regulates COPD through downstream targets has not been reported. The gratifying discovery in this study is that DFFB is a downstream target of miR-421. Previous studies have shown that DNA fragmentation factor B (DFFB) is closely related to cell apoptosis.41 However, its role in COPD cell apoptosis had not been reported yet. In this study, we found that CSE significantly increased the expression of DFFB, and miR-421 participated in regulating the apoptosis of HPMECs and HBEC in COPD induced by CSE through negative regulation of DFFB expression.
Several limitations need to be noted regarding the present study. The main limitation of this study is that in vitro CSE treatment only simulates a small fraction of the actual conditions that occur in COPD. In the future, it is necessary to construct in vivo animal COPD models and further study the regulatory mechanism of the MEG3/miR-421/DFFB signal axis in COPD.
In summary, this study demonstrates that MEG3 may participate in the process of COPD by regulating the miR-421/DFFB axis, and may be closely related to the apoptosis of HPMECs and HBEC. These results provide clues for understanding the possible mechanisms of treating COPD and are expected to provide potential therapeutic targets for COPD.
Data Sharing Statement
All data generated or analyzed during this study are included in this published article.
Funding
This study is supported by the Basic Research Project of Changzhou Science and Technology Bureau (CJ20200107).
Disclosure
The authors declare that they have no conflicts of interest.
References
1. Wu YX, Zuo YH, Cheng QJ, et al. Respiratory aspergillus colonization was associated with relapse of acute exacerbation in patients with chronic obstructive pulmonary disease: analysis of data from a retrospective cohort study. Front Med. 2021;8:640289. doi:10.3389/fmed.2021.640289
2. Trigueros JA, Garin N, Baloira A, et al. Cost-effectiveness analysis of triple therapy with budesonide/ glycopyrronium/ formoterol fumarate versus dual therapy in patients with chronic obstructive pulmonary disease in Spain. Int J Chron Obstruct Pulmon Dis. 2022;17:2905–2917. doi:10.2147/COPD.S384591
3. Hu J, Gao R, Wang Y, et al. Effect of Liuzijue on pulmonary rehabilitation in patients with chronic obstructive pulmonary disease: study protocol for a multicenter, non-randomized, prospective study. BMC Complement Med Ther. 2022;22(1):296. doi:10.1186/s12906-022-03789-6
4. Salvi S. Tobacco smoking and environmental risk factors for chronic obstructive pulmonary disease. Clin Chest Med. 2014;35(1):17–27. doi:10.1016/j.ccm.2013.09.011
5. Liang Y, Chang C, Zhu H, Shen N, He B, Yao W. Correlation between decrease of CRP and resolution of airway inflammatory response, improvement of health status, and clinical outcomes during severe acute exacerbation of chronic obstructive pulmonary disease. Intern Emerg Med. 2015;10(6):685–691. doi:10.1007/s11739-015-1228-2
6. Bi H, Wang G, Li Z, et al. Long noncoding RNA (lncRNA) maternally expressed gene 3 (MEG3) participates in chronic obstructive pulmonary disease through regulating human pulmonary microvascular endothelial cell apoptosis. Med Sci Monit. 2020;26:e920793. doi:10.12659/MSM.920793
7. Sun Y, An N, Li J, et al. miRNA-206 regulates human pulmonary microvascular endothelial cell apoptosis via targeting in chronic obstructive pulmonary disease. J Cell Biochem. 2019;120(4):6223–6236. doi:10.1002/jcb.27910
8. MacNee W. Accelerated lung aging: a novel pathogenic mechanism of chronic obstructive pulmonary disease (COPD). Biochem Soc Trans. 2009;37(4):819–823. doi:10.1042/bst0370819
9. Zhang H, Sun D, Li D, et al. Long non-coding RNA expression patterns in lung tissues of chronic cigarette smoke induced COPD mouse model. Sci Rep. 2018;8(1):7609. doi:10.1038/s41598-018-25702-3
10. Zong L, Wang Q, Song J. LncRNA MIR155HG contributes to smoke-related chronic obstructive pulmonary disease by targeting miR-128-5p/BRD4 axis. Biosci Rep. 2020;40(3). doi:10.1042/bsr20192567
11. Mei J, Zhang Y, Lu S, Wang J. Long non-coding RNA NNT-AS1 regulates proliferation, apoptosis, inflammation and airway remodeling of chronic obstructive pulmonary disease via targeting miR-582-5p/FBXO11 axis. Biomed Pharmacother. 2020;129:110326. doi:10.1016/j.biopha.2020.110326
12. Luo J, Li L, Hu D, Zhang X. LINC00612/miR-31-5p/Notch1 axis regulates apoptosis, inflammation, and oxidative stress in human pulmonary microvascular endothelial cells induced by cigarette smoke extract. Int J Chron Obstruct Pulmon Dis. 2020;15:2049–2060. doi:10.2147/COPD.S255696
13. Gu W, Yuan Y, Wang L, et al. Long non-coding RNA TUG1 promotes airway remodelling by suppressing the miR-145-5p/DUSP6 axis in cigarette smoke-induced COPD. J Cell Mol Med. 2019;23(11):7200–7209. doi:10.1111/jcmm.14389
14. Fan S, Ren Y, Zhang W, Zhang H, Wang C. Long non-coding maternally expressed gene 3 regulates cigarette smoke extract-induced apoptosis, inflammation and cytotoxicity by sponging miR-181a-2-3p in 16HBE cells. Oncol Lett. 2021;21(1):45. doi:10.3892/ol.2020.12306
15. Song B, Ye L, Wu S, Jing Z. Long non-coding RNA MEG3 regulates CSE-induced apoptosis and inflammation via regulating miR-218 in 16HBE cells. Biochem Biophys Res Commun. 2020;521(2):368–374. doi:10.1016/j.bbrc.2019.10.135
16. Piletic K, Kunej T. MicroRNA epigenetic signatures in human disease. Arch Toxicol. 2016;90(10):2405–2419. doi:10.1007/s00204-016-1815-7
17. Xue H, Li MX. MicroRNA-150 protects against cigarette smoke-induced lung inflammation and airway epithelial cell apoptosis through repressing p53: microRNA-150 in CS-induced lung inflammation. Hum Exp Toxicol. 2018;37(9):920–928. doi:10.1177/0960327117741749
18. Dang X, Yang L, Guo J, et al. miR-145-5p is associated with smoke-related chronic obstructive pulmonary disease via targeting KLF5. Chem Biol Interact. 2019;300:82–90. doi:10.1016/j.cbi.2019.01.011
19. An J, Yang T, Dong J, et al. Identifying miRNA modules and related pathways of chronic obstructive pulmonary disease associated emphysema by weighted gene co-expression network analysis. Int J Chron Obstruct Pulmon Dis. 2021;16:3119–3130. doi:10.2147/COPD.S325300
20. Han DSC, Ni M, Chan RWY, et al. The biology of cell-free DNA fragmentation and the roles of DNASE1, DNASE1L3, and DFFB. Am J Hum Genet. 2020;106(2):202–214. doi:10.1016/j.ajhg.2020.01.008
21. Sohn SH, Kim KN, Kim IK, Lee EI, Ryu JJ, Kim MK. Effects of tobacco compounds on gene expression in fetal lung fibroblasts. Environ Toxicol. 2008;23(4):423–434. doi:10.1002/tox.20335
22. Peng Y, Cheng W, Duan J, et al. Prohibitin protects pulmonary microvascular endothelial cells against cigarette smoke extract-induced cell apoptosis and inflammation. Int J Chron Obstruct Pulmon Dis. 2022;17:653–665. doi:10.2147/COPD.S345058
23. Mastalerz M, Dick E, Chakraborty A, et al. Validation of in vitro models for smoke exposure of primary human bronchial epithelial cells. Am J Physiol Lung Cell Mol Physiol. 2022;322(1):L129–L148. doi:10.1152/ajplung.00091.2021
24. Zhou AY, Zhao YY, Zhou ZJ, et al. Microarray analysis of long non-coding RNAs in lung tissues of patients with COPD and HOXA-AS2 promotes HPMECs proliferation via Notch1. Int J Chron Obstruct Pulmon Dis. 2020;15:2449–2460. doi:10.2147/COPD.S259601
25. Wu X. Depletion of miR-380 mitigates human bronchial epithelial cells injury to improve chronic obstructive pulmonary disease through targeting CHRNA4. Mol Cell Probes. 2020;49:101492. doi:10.1016/j.mcp.2019.101492
26. Decramer M, Janssens W, Miravitlles M. Chronic obstructive pulmonary disease. Lancet. 2012;379(9823):1341–1351. doi:10.1016/s0140-6736(11)60968-9
27. Hutchinson D, Muller J, McCarthy JE, et al. Cadmium nanoparticles citrullinate cytokeratins within lung epithelial cells: cadmium as a potential cause of citrullination in chronic obstructive pulmonary disease. Int J Chron Obstruct Pulmon Dis. 2018;13:441–449. doi:10.2147/COPD.S152028
28. Zhuo SM, Li SC, Lin YQ, Yu HB, Li N. The effects of anti-Fas ribozyme on T lymphocyte apoptosis in mice model with chronic obstructive pulmonary disease. Iran J Basic Med Sci. 2017;20(10):1102–1108. doi:10.22038/IJBMS.2017.9367
29. Loeckx M, Rabinovich RA, Demeyer H, et al. Smartphone-based physical activity telecoaching in chronic obstructive pulmonary disease: mixed-methods study on patient experiences and lessons for implementation. JMIR mHealth uHealth. 2018;6(12):e200. doi:10.2196/mhealth.9774
30. Wang J, Xu W, He Y, Xia Q, Liu S. LncRNA MEG3 impacts proliferation, invasion, and migration of ovarian cancer cells through regulating PTEN. Inflamm Res. 2018;67(11–12):927–936. doi:10.1007/s00011-018-1186-z
31. Tong P, Peng QH, Gu LM, Xie WW, Li WJ. LncRNA-MEG3 alleviates high glucose induced inflammation and apoptosis of retina epithelial cells via regulating miR-34a/SIRT1 axis. Exp Mol Pathol. 2019;107:102–109. doi:10.1016/j.yexmp.2018.12.003
32. Zhang W, Shi S, Jiang J, Li X, Lu H, Ren F. LncRNA MEG3 inhibits cell epithelial-mesenchymal transition by sponging miR-421 targeting E-cadherin in breast cancer. Biomed Pharmacother. 2017;91:312–319. doi:10.1016/j.biopha.2017.04.085
33. Huang H, Zheng S, Lu M. Downregulation of lncRNA MEG3 is involved in Parkinson’s disease. Metab Brain Dis. 2021;36(8):2323–2328. doi:10.1007/s11011-021-00835-z
34. Liu Y, Zhuang Y, Fu X, Li C. LncRNA POU3F3 promotes melanoma cell proliferation by downregulating lncRNA MEG3. Discov Oncol. 2021;12(1):21. doi:10.1007/s12672-021-00414-9
35. Mohammed SR, Shaker OG, Mohamed MM, et al. The emerging role of lncRNA MEG3 and MEG3 rs7158663 in hepatocellular carcinoma. Eur Rev Med Pharmacol Sci. 2022;26(1):11–21. doi:10.26355/eurrev_202201_27742
36. Lei Z, Guo H, Zou S, Jiang J, Kui Y, Song J. Long non-coding RNA maternally expressed gene regulates cigarette smoke extract induced lung inflammation and human bronchial epithelial apoptosis via miR-149-3p. Exp Ther Med. 2021;21(1):60. doi:10.3892/etm.2020.9492
37. Momen-Heravi F, Bala S. Emerging role of non-coding RNA in oral cancer. Cell Signal. 2018;42:134–143. doi:10.1016/j.cellsig.2017.10.009
38. Shen Q, Zheng J, Wang X, Hu W, Jiang Y, Jiang Y. LncRNA SNHG5 regulates cell apoptosis and inflammation by miR-132/PTEN axis in COPD. Biomed Pharmacother. 2020;126:110016. doi:10.1016/j.biopha.2020.110016
39. Bartel DP. MicroRNAs: target recognition and regulatory functions. Cell. 2009;136(2):215–233. doi:10.1016/j.cell.2009.01.002
40. Diao X, Zhou J, Wang S, Ma X. Upregulation of miR-132 contributes to the pathophysiology of COPD via targeting SOCS5. Exp Mol Pathol. 2018;105(3):285–292. doi:10.1016/j.yexmp.2018.10.002
41. McDonald JM, Dunmire V, Taylor E, et al. Attenuated expression of DFFB is a hallmark of oligodendrogliomas with 1p-allelic loss. Mol Cancer. 2005;4:35. doi:10.1186/1476-4598-4-35
 © 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Downregulation of Lysosome-Associated Membrane Protein-2A Contributes to the Pathogenesis of COPD
Jeong YJ, Lee KH, Woo J, Kim JY, Lee CH, Yoo CG
International Journal of Chronic Obstructive Pulmonary Disease 2023, 18:289-303
Published Date: 14 March 2023
LncRNA RP11-521C20.3 Inhibits Cigarette Smoke Extract-Induced Apoptosis in A549 Cells by Targeting BMF Signaling
Zhong Y, Li C, Xiang Y, Zhou J, Zhang J
International Journal of Chronic Obstructive Pulmonary Disease 2023, 18:669-682
Published Date: 21 April 2023
Silencing KLF6 Alleviates Cigarette Smoke Extract-Induced Mitochondrial Dysfunction in Bronchial Epithelial Cells by SIRT4 Upregulation
Wan M, Wang C, Cui J, Xia Q, Zhang L
International Journal of Chronic Obstructive Pulmonary Disease 2024, 19:815-828
Published Date: 23 March 2024
A miRNA-21-Mediated PTEN/Akt/NF-κB Axis Promotes Chronic Obstructive Pulmonary Disease Pathogenesis
Sai X, Qin C, Zhang Z, Yu H, Bian T
International Journal of Chronic Obstructive Pulmonary Disease 2024, 19:1141-1151
Published Date: 25 May 2024