")
Back to Journals » Infection and Drug Resistance » Volume 17
Prevalence and Molecular Characteristics of Enterococci Isolated from Clinical Bovine Mastitis Cases in Ningxia
Authors Liu J , Liang Z , Zhongla M, Wang H, Sun X , Zheng J, Ding X, Yang F
Received 26 January 2024
Accepted for publication 25 April 2024
Published 27 May 2024 Volume 2024:17 Pages 2121—2129
DOI https://doi.org/10.2147/IDR.S461587
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Suresh Antony
Jing Liu,1,* Zeyi Liang,1,* Maocao Zhongla,2 Hongsheng Wang,3 Xu Sun,4 Juanshan Zheng,1 Xuezhi Ding,1 Feng Yang1
1Key Laboratory of New Animal Drug Project of Gansu Province/Key Laboratory of Veterinary Pharmaceutics Discovery, Lanzhou Institute of Husbandry and Pharmaceutical Sciences of Chinese Academy of Agricultural Science, Lanzhou, People’s Republic of China; 2Gannan Animal Disease Prevention and Control Center, Hezuo, People’s Republic of China; 3Xiangyang Vocational and Technical College, Xiangyang, People’s Republic of China; 4College of Life Sciences and Engineering, Lanzhou University of Technology, Lanzhou, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Feng Yang; Xuezhi Ding, Lanzhou Institute of Husbandry and Pharmaceutical Sciences of Chinese Academy of Agricultural Science, No. 335 Jiangouyan, Qilihe District, Lanzhou, Gansu, 730050, People’s Republic of China, Tel +86-931-2115262, Fax +86-931-2114180, Email [email protected]; [email protected]
Purpose: This study aimed to investigate the prevalence and genetic characterization of enterococcal isolates (Enterococcus faecalis, Enterococcus faecium and Enterococcus hirae) isolated from clinical bovine mastitis cases in Ningxia, China.
Patients and Methods: The enterococci were identified by 16S rRNA amplification and sequencing. Antimicrobial resistance was determined by disc diffusion method. Virulence and antimicrobial resistance genes were detected by PCR assays.
Results: Overall, 198 enterococcal isolates were identified from 2897 mastitis samples, including 137 (4.7%) E. faecalis, 50 (1.7%) E. faecium and 11 (0.4%) E. hirae. E. faecalis, E. faecium and E. hirae isolates showed high resistance to tetracycline (92.7%, 68.0%, 90.9%), followed by erythromycin (86.9%, 76.0%, 72.7%). The multidrug-resistant strains of E. faecalis and E. faecium were 29 (21.2%) and 13 (26.0%), respectively. The resistance of E. faecalis, E. faecium and E. hirae isolates to tetracycline is mainly attributed to the presence of tetL (alone or combined with tetM and/or tetK), the erythromycin resistance to ermB (alone or combined with ermC and/or ermA). Moreover, cpd (94.2%), gelE (77.4%), efaAfs (93.4%), and esp (79.6%) were the most common virulence genes in E. faecalis. In E. faecium, except for the gene efaAfs (82.0%), other virulence genes are rarely found. Only two strains of E. hirae carrying asa1 gene were detected.
Conclusion: The results of this study can provide a reference for the prevention and treatment of bovine mastitis caused by enterococci.
Keywords: bovine mastitis, enterococci, antimicrobial resistance, virulence
A Letter to the Editor has been published for this article.
Introduction
Mastitis is the most common disease of bovines, and it adversely affects animal health and the profitability of dairy farms, including increases in bovine mortality and decreases in milk production.1 Greater progress has been made in controlling infectious mastitis pathogens through improved milking room hygiene and feeding management, but for opportunistic mastitis pathogens present in the environment, such as enterococci, these management measures are less effective. Enterococci is a facultative anaerobic gram-positive bacteria found in intestines of humans and animals. According to the consistency of 16S rRNA sequence, enterococci can be divided into several subgroups, such as E. faecalis, E. faecium, E. hirae. They are highly tolerant to the environment and have a variety of host niches. Several studies in the past have shown that enterococci can cause mastitis in dairy bovines.2–4
At present, antibiotics are still the main treatment for bovine mastitis. However, this method can lead to an increased incidence of antimicrobial resistant infections in bovines during long-term rearing, increasing production costs and even causing public health incidents. Enterococci could develop resistance to most existing antibiotics by mutation or incorporation of foreign genetic material,5 and it may be possible for these antibiotic resistances to be transmitted in other bacteria by exchange of genetic information (plasmids and transposon) through mobile genetic elements.6 Previous studies have reported that E. faecium isolated from food is resistant to a variety of antibiotics, including aminoglycosides, macrolide, β-lactam, and tetracycline.7 Recent studies have attributed enterococci toxicity to various factors, such as gelatinases, surface proteins, sex pheromones, collagen-binding cell wall proteins, polymeric substances, and biofilm formation.2
At present, enterococci have been reported worldwide. Due to the diversity and variability of enterococci genes,8 a deeper understanding of the genotype of enterococci and its epidemiology in dairy bovines may help to monitor the emergence of highly virulent strains or antibiotic resistant strains. The purpose of this study was to study the prevalence, antimicrobial resistance, and virulence characteristics of predominant enterococci (E. faecalis, E. faecium and E. hirae) isolated from clinical bovine mastitis in Ningxia Hui Autonomous Region of China.
Materials and Methods
Sample Collection and Enterococci Isolates
Mastitis milk samples of dairy cows (n=2897) were collected from 21 commercial dairy herds in Ningxia Hui Autonomous Region, China, from July 2018 to May 2023. Clinical mastitis cases were confirmed by the abnormal signs of bovine udder (oedema, lumps, increase in temperature, hardening or pain) and milk samples (presence of clots, flakes or blood).9 After transportation to the laboratory, 100 μL of each sample was plated on sheep blood agar (Huan kai, Guangdong, China) at 37 °C aerobically for 18 to 24 h. Colonies were initially identified as enterococcal isolates based on the morphological features (shape, color, and size).10–13 The assumed isolates were further confirmed by PCR and sequencing as described in our previous study.9 Briefly, the genomic DNA was extracted by the Bacterial DNA Kit (Qiagen, Hilden, Germany) following the manufacturer’s protocol. The 16S rRNA gene was amplified by the 16S rDNA Bacterial Identification PCR Kit (Takara, Shiga, Japan) according the manufacturer’s recommendation. The PCR products were purified and sequenced through Sanger sequencing (Tsingke, Xi’an, China). BLAST analysis tool from NCBI (https://blast.ncbi.nlm.nih.gov/Blast.cgi) was used to analyze the nucleotide sequences obtained.
Antimicrobial Susceptibility Testing
The antimicrobial susceptibility of enterococcal isolates to 8 antimicrobial agents was determined using the disk diffusion method on Mueller Hinton agar (Oxoid, Basingstoke, UK) plates according to the standards of the Clinical and Laboratory Standards Institute (CLSI, 2018). The (Oxoid) antibiotic discs included tetracycline (30 μg), vancomycin (30 μg), penicillin (10 μg), quinupristin (15 μg), erythromycin (15 μg), ciprofloxacin (5 μg), chloramphenicol (30 μg), and rifampin (5 μg). Multidrug-resistant (MDR) isolates were defined as an isolate being resistant to at least 3 antimicrobial agents belonging to different antimicrobial categories (Magiorakos et al, 2012). Staphylococcus aureus ATCC 25923 was used as quality control strains.
Genetic Determinants for Antimicrobial Resistance and Virulence
Resistance genes and virulence genes were detected by PCR, and primers and thermal conditions were summarized in Table S1. The resistance-associated genes for tetracycline (tetK, tetL, tetM, tetO and tetS) and erythromycin (ermA, ermB, and ermC) were searched using PCR as previously described.14–19 The virulence factors sex pheromone determinant (cpd), cytolysin (cylA), gelatinase (gelE), regulator of the expression of gelE (frs), hyaluronidase (hyl), cell wall adhesins (efaAfs, efaAfm, efaA), aggregation substance (asa1), enterococcal surface protein (esp), collagen adhesion (ace) were also evaluated through PCR.20–22 PCR products (10 μL) were analyzed by electrophoresis on 1.2% agarose gel and photographed using a UV imaging system.
Results
Antimicrobial Susceptibility of Enterococci
In this study, a total of 198 enterococcal isolates (6.8%) were isolated from 2897 clinical mastitic samples, including E. faecalis (137, 4.7%), E. faecium 50 (1.7%) and E. hirae 11 (0.4%). The antimicrobial susceptibility of these isolates to 8 antibiotics was evaluated and summarized in Table 1. The results showed that 171 (86.4%) enterococcal isolates were resistant to tetracycline (92.7% for E. faecalis; 68.0% for E. faecium; 90.9% for E. hirae) and 166 (83.8%) to erythromycin (86.9% for E. faecalis; 76.0% for E. faecium; 72.7% for E. hirae). Resistance against rifampin and chloramphenicol was found in 45 (22.7%) and 23 (11.6%) isolates, respectively. Quinupristin-dalfopristin, ciprofloxacin and penicillin resistance were measured in 30 (15.2%), 24 (12.1%) and 10 (5.1%) of the enterococcal isolates except for E. hirae, respectively. None of the isolates showed resistance to vancomycin. In addition, multidrug resistance was found in 42 (21.2%) enterococcal isolates, including E. faecalis (29, 21.2%) and E. faecium (13, 26.0%).
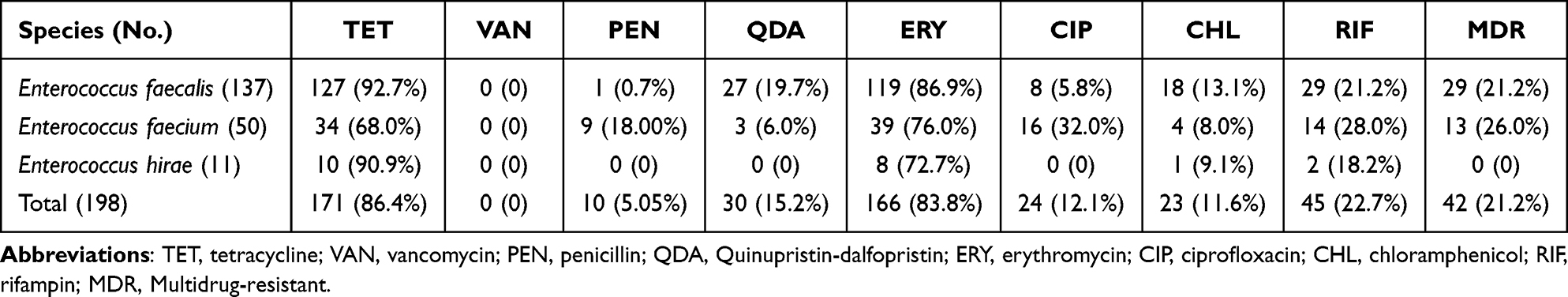 |
Table 1 Antimicrobial Resistance Characteristics of Enterococci Isolates |
Genotypic Resistance Profiles of Enterococci
In this study, the enterococcal isolates showed higher resistance to tetracycline and erythromycin in comparison with other tested antimicrobial agents. Therefore, we tested the antimicrobial resistance gene frequencies of tetracycline (tetK, tetL, tetM, tetO, and tetS) and erythromycin (ermA, ermB, and ermC) (Table 2). The tetracycline-resistant gene tetL was detectd in 196 (99.0%) enterococcal isolates, followed by tetM (170, 85.9%), tetK (118, 59.6%), tetS (90, 45.5%) and tetO (18, 9.1%). One hundred and ninety-six tetL alone or combined with tetM, tetK, tetS and/or tetO was found in all tetracycline-resistant isolates. However, 25 tetL-carrying (alone or combined with tetM, tetK, tetL, tetS and/or tetO) isolates, including 10 E. faecalis, 14 E. faecium, and 1 E. hirae, were susceptible to tetracycline. Additionally, ermB, ermC and ermA were found in 189 (95.5%), 14 (7.1%) and 2 (1.0%) enterococcal isolates, respectively. One hundred and ninety ermB alone or combined with ermC and/or ermA was found in all erythromycin-resistant isolates. However, 23 ermB-carrying (alone or combined with ermC and/or ermA) isolates, including 10 E. faecalis, 11 E. faecium, and 3 E. hirae, were susceptible to erythromycin.
 |
Table 2 Characterization of Drug Resistance Genes in Enterococci Isolates |
Genotypic Virulence Profiles of Enterococci
The presence of virulent genes in enterococcal isolates are shown in Table 3. E. faecalis isolates carried most of the tested genes, including cpd (129, 94.2%), efaAfs (128, 93.4%), esp (109, 79.6%), gelE (106, 77.4%), ace (43, 31.6%), efaAfm (38, 27.7%), asa1 (38, 27.7%) and cylA (22, 16.1%). In E. faecium, the efaAfs (41, 82.0%) was the most prevalent virulence gene, followed by asa1 (8, 16.0%), efaAfm (3, 6.0%), cpd (2, 4.0%), esp (2, 4.0%), and ace (2, 4.0%). However, aside from 2 asa1-positive E. hirae isolates, none of the other virulent genes were detected in this species.
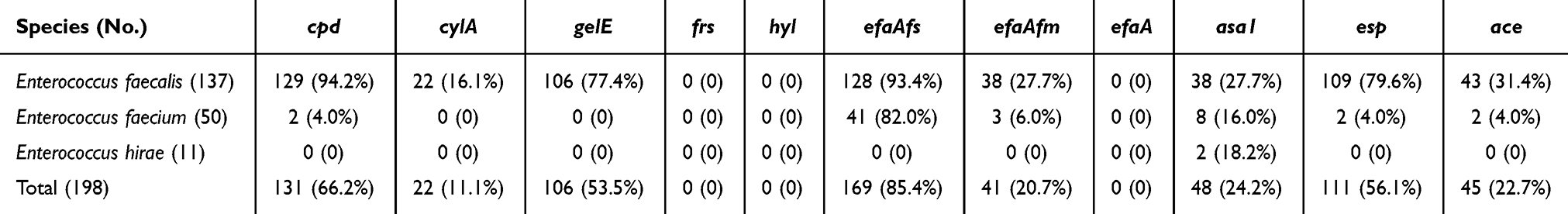 |
Table 3 Characterization of Virulence Genes in Enterococci Isolates |
Discussion
Enterococci are the main environmental pathogens causing bovine mastitis and antimicrobials are widely used for the treatment of these infections.23,24 Unfortunately, enterococcus has inherent resistance to many broad-spectrum antibiotics, posing a significant threat to public health.25 Understanding the prevalence, antimicrobial resistance and virulence factors of enterococci is of great value for targeted therapy and control of bovine mastitis. In the present study, E. faecalis, E. faecium, and E. hirae were detected in 4.7%, 1.7%, and 0.4% of 2897 mastitic samples, respectively. This is a slight increase from the prevalence of E. faecalis (4.5%) reported in previous studies in China.2 It is lower than rates reported in California, USA,26 but higher than the E. faecalis (0.2%), E. faecium (0.2%) reported in Brandenburg, Germany.27 The management system of the farm, source of samples, climate, and herd size may explain some differences in the research results.28 As reported in the past, the prevalence of E. hirae is much lower than that of E. faecalis and E. faecium.29,30 It is worth noting that recent research has shown that E. hirae is a cause of infection in various animal diseases.31 At present, reports of E. hirae are limited worldwide, resulting in a lack of comprehensive reports on clinical features and management.32 This study supplements the clinical characteristics of E. faecalis, E. faecium, and E. hirae, which causes mastitis in northwest China.
Antibiotic therapy has been used as one of the successful strategies to control mastitis caused by enterococci.33 The β-Lactam, tetracyclines, macrolides and aminoglycosides are commonly used to treat bovine mastitis.34 Unfortunately, the increase in the number of drug-resistant strains hinders the effectiveness of antibacterial treatment.35 It is worth noting that previous studies have reported that E. faecalis is prone to acquire antimicrobial resistance.15 Our study found that E. faecalis, E. faecium and E. hirae showed high resistance to tetracycline and erythromycin. These data are similar to those contained in other reports.2,36–38 In addition, previous studies have also reported high resistance to tetracycline and erythromycin in E. faecalis isolated from food animals.15,39 The reason of this result may be due to the long-term and widespread use of tetracycline and erythromycin in non-targeted therapies.2,40 Thus, antimicrobials such as tetracycline and erythromycin should be used cautiously when treating cow mastitis caused by enterococci in Ningxia, China. In the present survey, the E. faecium and E. hirae isolates displayed fewer resistance phenotypes than the E. faecalis. This is consistent with previous research results.41 Besides, our results indicate that E. faecalis and E. faecalis have relatively low resistance rates to vancomycin, quinupristin-dalfopristin, ciprofloxacin, chloramphenicol, and rifampicin. The lower proportion of antibiotic resistance may be due to the lower frequency of use of these antibiotics in dairy farms. However, the multidrug resistant strains were still observed in 21.2% and 26.0% of the E. faecalis and E. faecium, respectively. This result suggests that we should use antibiotics with caution. Because this may be due to increasing exposure to antimicrobial environments can lead to an increase in the number of multidrug-resistant strains. Therefore, the frequent use of these antibiotics in the treatment of mastitis or other diseases may explain the high phenotype of multidrug resistance. Previous studies have shown that enterococci is a common multidrug resistant bacterium.42,43 Enterococci may transfer resistance genes to more pathogenic bacteria through mobile genetic elements (MGE).44 Therefore, these resistant agricultural isolates may be important stepping stones in the evolution of enterococci into resistant human pathogens.45
In this study, the most commonly antimicrobial resistance of the three enterococcal species identified was resistance to erythromycin and tetracycline. Therefore, this study tested the genes that develop the corresponding resistance. The resistance of enterococci to tetracyclines is mainly mediated by the tet genes. In this study, the tetL and tetM gene was the most prevalent gene among the tetracycline-resistant isolates. The tetL and tetM gene has been found to be widespread among enterococci and is often associated with high-level tetracycline resistance.46 In addition, the tet genes are usually carried on plasmids or transposons, which can easily spread between bacteria, leading to the dissemination of antibiotic resistance.47 The resistance of enterococci to macrolides is mainly mediated by the erm genes.48 The erm gene modifies the target site on the bacterial ribosome to prevent the binding of the antibiotics, thereby producing a high level of resistance to macrolides.26 In this study, the ermB gene was the most prevalent gene among the erythromycin-resistant isolates. The ermB genes encode methylases, which induces N6-dimethylation at A2071 of the 23S rRNA,49 leading to the inhibition of macrolide binding.50 The erm genes are usually carried on conjugative transposons, which can move between bacteria and spread antibiotic resistance.51 The results of antibiotics resistance gene testing in this study were supported by previous studies, and tetL, tetM, and ermB were more commonly observed in enterococci.36,52,53 It is noteworthy that 25 isolates carrying at least one tet (tetK, tetL, tetM, tetO, and tetS) gene are sensitive to tetracycline, and 23 isolates carrying at least one erm (ermA, ermB, and ermC) gene are sensitive to erythromycin. This difference can be attributed to the lack of expression of these resistance genes.54
Recent studies have proved that enterococci from food of animal origin may be related to the colonization and pathogenesis of human diseases.22 This requires extensive and sustained attention to the encoding and production of multiple virulence factors by potentially pathogenic enterococci. In the current study, the E. faecalis isolates exhibited high prevalence of cpd, efaAfs, esp and gelE, and the efaAfs was the most prevalent virulence gene in E. faecium. Similar observations have also been reported by other reports.2,22 The first step of enterococci infection is colonization in tissues, which is mainly mediated by adhesins anchored by cell walls.55 In this study, six adhesive virulence genes (efaAfs, efaAfm, efaA, esp, ace, asa1) were detected, except for efaA gene, other genes were detected in E. faecalis and E. faecium. In this study, efaAfs, efaAfm, esp, and ace genes were negative in all of E. hirae. Recent study also failed to observe efaAfs and efaAfm genes in E. hirae isolates.56 However, another study observed efaAfs and efaAfm genes in important foodborne E. hirae.57 This difference may be due to the low prevalence of toxins producing E. hirae isolates in the study area. In addition, E. hirae may have other untested toxin genes related to bacterial adhesion.58 The efaAfm and efaAfs genes are believed to be involved in adhesion to host cells and biofilm formation.59 The efaAfm and efaAfs genes were present in a high proportion of E. faecalis and E. faecium isolates. This study found that 109 strains of E. faecalis and 2 strains of E. faecium carry the esp gene. Previous studies have found esp genes contribute to the establishment of successful enterococci infections and increase the colonization and persistence of infections.60,61 This study found that 43 strains of E. faecalis and 2 strains of E. faecium carry the ace gene. The ace gene encodes a collagen adhesin. When the host cells are damaged, the adhesion proteins are exposed, which will promote the adhesion of bacteria to the host cells. Studies have shown that enterococci carrying ace gene has stronger adhesion ability.60 This study found that 38 strains of E. faecalis, 8 strains of E. faecium, and 2 strains of E. hirae carry the asa1 gene. The asa1 gene encodes a polymer that binds to extracellular matrix proteins such as fibronectin, thrombospondin, vitronectin, and type I collagen and mediates multiple stages of infection.62,63 Therefore, we speculate that isolates of E. faecalis and E. faecalis may have strong initial colonization and biofilm formation abilities.
The cpd gene encodes a sex pheromone, a sex pheromone thought to be involved in triggering an inflammatory response.8 The sex pheromones secreted by bacteria are perceived by potential companion cells, which mediate the contact between bacteria and induce the lateral transfer of the coupling plasmid encoding the determinants of antibiotic resistance and virulence.64,65 Similar to the results of the present study, cpd gene was also found to have the higher detection rate in E. faecalis in past studies.22 However, only two strains of E. faecalis were found to carry the cpd gene, and all strains of E. hirae were negative for the cpd gene.
The production of cytolysin has been proved by many experiments to be related to the severity of diseases induced by enterococci.66,67 Cytolysin has a wide range of target cells, so it is also one of the virulence traits that attract much attention in pathogenic bacteria.68,69 In this study, we observed the cylA gene associated with hemolysin in 16.1% of E. faecalis isolates. This result is higher than previous research results.53,70 The cylA gene synthesizes serine protease, which causes the activation of cytolysin outside the cell.71 This reaction may cause red blood cell lysis and tissue damage, leading to bacterial transmission within the host and promoting the development of infection. In addition, cytolysin can escape the host immune system by invading and avoiding phagocyte killing.72
In addition, we evaluated the gelatinase-related toxin genes. The gelE gene was present in 77.4% of E. faecalis. This agrees with the results of previous studies on isolated strains of E. faecium, they also did not detect the gelE gene in E. faecium.8,71,73 Encoded by the chromosomal gene gelE, gelatinase is an extracellular zinc endopeptidase that hydrolyzes a variety of proteins or peptides, including casein, hemoglobin, insulin, fibrinogen and collagen.73 The decomposition of these proteins may contribute to the invasion and spread of pathogenic bacteria to the breast.74 Therefore, gelatinase is considered to affect the virulence of enterococci and is related to the biofilm formation, infection and spread of enterococci.75,76 The expression of the gelE gene is regulated by a quorum sensing system, which allows the bacterium to coordinate its behavior with other members of its population.77 When the bacteria reach a certain population density, the quorum sensing system is activated, which leads to the expression of virulence factors including gelatinase.
Conclusion
This study provided rich species diversity for enterococci causing mastitis in Ningxia. It is worth noting that although the incidence rate of E. faecalis is relatively low in clinical bovine mastitis cases, it has an upward trend compared with previous studies. The E. faecalis, E. faecium, and E. hirae displayed high phenotypic and genotypic resistance to tetracycline and erythromycin, which requires more attention through comprehensive monitoring of the antimicrobials use in dairy production. In addition, the high prevalence of cpd, gelE, efaAfs, and esp in E. faecalis suggests that they may play an important role in causing clinical bovine mastitis in China. Because of the complexity of clinical bovine mastitis caused by enterococci, additional measures must be taken to reduce the development and spread of antimicrobial resistant elements from farms to clinics around the world. In addition, although E. hirae carries fewer toxin genes in this study, more extensive studies are needed to prove the true infectivity and pathogenicity of E. hirae in mastitis.
Ethics Approval and Informed Consent
The animal owners were informed about the purpose of the study and consent of each animal owner was obtained before the physical examination of cows for clinical mastitis and the collection of milk samples. All procedures involved in animal care and their use were in strict accordance with the guidelines for the Care and Use of Laboratory Animals, Lanzhou Institute of Husbandry and Pharmaceutical Sciences, CAAS, China (SYXK-2019-0012).
Acknowledgments
This work was supported by the National Key R&D Program of China during the 14th Five year Plan Period (Grant No. 2022YFD1302101) and the National Natural Science Foundation of China (Grant No. 32350410415).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Hertl J, Schukken Y, Bar D, et al. The effect of recurrent episodes of clinical mastitis caused by gram-positive and gram-negative bacteria and other organisms on mortality and culling in Holstein dairy cows. J Dairy Sci. 2011;94(10):4863–4877. doi:10.3168/jds.2010-4000
2. Yang F, Zhang S, Shang X, et al. Antimicrobial resistance and virulence genes of Enterococcus faecalis isolated from subclinical bovine mastitis cases in China. J Dairy Sci. 2019;102(1):140–144. doi:10.3168/jds.2018-14576
3. Odierno L, Calvinho L, Traverssa P, Lasagno M, Bogni C, Reinoso E. Conventional identification of Streptococcus uberis isolated from bovine mastitis in Argentinean dairy herds. J Dairy Sci. 2006;89(10):3886–3890. doi:10.3168/jds.S0022-0302(06)72431-6
4. Nonnemann B, Lyhs U, Svennesen L, Kristensen KA, Klaas IC, Pedersen K. Bovine mastitis bacteria resolved by MALDI-TOF mass spectrometry. J Dairy Sci. 2019;102(3):2515–2524. doi:10.3168/jds.2018-15424
5. García-Solache M, Rice LB. The enterococcus: a model of adaptability to its environment. Clin Microbiol Rev. 2019;32(2):e00058–18. doi:10.1128/CMR.00058-18
6. Nasiri M, Hanifian S. Enterococcus faecalis and Enterococcus faecium in pasteurized milk: prevalence, genotyping, and characterization of virulence traits. LWT. 2022;153:112452. doi:10.1016/j.lwt.2021.112452
7. Daza MVB, Milani G, Cortimiglia C, Pietta E, Bassi D, Cocconcelli PS. Genomic insights of Enterococcus faecium UC7251, a multi-drug resistant strain from ready-to-eat food, highlight the risk of antimicrobial resistance in the food chain. Front Microbiol. 2022;13:894241.
8. Eaton TJ, Gasson MJ. Molecular screening of Enterococcus virulence determinants and potential for genetic exchange between food and medical isolates. Appl Environ Microbiol. 2001;67(4):1628–1635. doi:10.1128/AEM.67.4.1628-1635.2001
9. Yang F, Shi W, Meng N, Zhao Y, Ding X, Li Q. Antimicrobial resistance and virulence profiles of staphylococci isolated from clinical bovine mastitis. Front Microbiol. 2023;14:1190790.
10. Margalho LP, van Schalkwijk S, Bachmann H, Sant’Ana AS. Enterococcus spp. in Brazilian artisanal cheeses: occurrence and assessment of phenotypic and safety properties of a large set of strains through the use of high throughput tools combined with multivariate statistics. Food Control. 2020;118:107425. doi:10.1016/j.foodcont.2020.107425
11. Fu M, Zhang X, Chen B, Li M, Zhang G, Cui L. Characteristics of isolates of Pseudomonas aeruginosa and Serratia marcescens associated with post-harvest Fuzi (Aconitum carmichaelii) rot and their novel loop-mediated isothermal amplification detection methods. Front Microbiol. 2021;12. doi:10.3389/fmicb.2021.705329
12. Tyc O, Zweers H, De Boer W, Garbeva P. Volatiles in inter-specific bacterial interactions. Front Microbiol. 2015;6:1412. doi:10.3389/fmicb.2015.01412
13. Delboni MG, Gomes BP, Francisco PA, Teixeira FB, Drake D. Diversity of Enterococcus faecalis genotypes from multiple oral sites associated with endodontic failure using repetitive sequence-based polymerase chain reaction and arbitrarily primed polymerase chain reaction. J Endodontics. 2017;43(3):377–382. doi:10.1016/j.joen.2016.10.042
14. Zhou N, Zhang J, Fan M, Wang J, Guo G, Wei X. Antibiotic resistance of lactic acid bacteria isolated from Chinese yogurts. J Dairy Sci. 2012;95(9):4775–4783. doi:10.3168/jds.2011-5271
15. Kim YB, Seo KW, Jeon HY, Lim S-K, Sung HW, Lee YJ. Molecular characterization of erythromycin and tetracycline-resistant Enterococcus faecalis isolated from retail chicken meats. Poultr Sci. 2019;98(2):977–983. doi:10.3382/ps/pey477
16. Shao Y, Zhang W, Guo H, Pan L, Zhang H, Sun T. Comparative studies on antibiotic resistance in Lactobacillus casei and Lactobacillus plantarum. Food Control. 2015;50:250–258. doi:10.1016/j.foodcont.2014.09.003
17. Aarestrup FM, Jensen LB. Trends in antimicrobial susceptibility in relation to antimicrobial usage and presence of resistance genes in Staphylococcus hyicus isolated from exudative epidermitis in pigs. Vet Microbiol. 2002;89(1):83–94. doi:10.1016/S0378-1135(02)00177-3
18. Feng Y, Qi W, Wang X-R, et al. Genetic characterization of antimicrobial resistance in Staphylococcus aureus isolated from bovine mastitis cases in Northwest China. J Integr Agric. 2016;15(12):2842–2847. doi:10.1016/S2095-3119(16)61368-0
19. Depardieu F, Perichon B, Courvalin P. Detection of the van alphabet and identification of enterococci and staphylococci at the species level by multiplex PCR. J Clin Microbiol. 2004;42(12):5857–5860. doi:10.1128/JCM.42.12.5857-5860.2004
20. Amaral DM, Silva LF, Casarotti SN, Nascimento LCS, Penna ALB. Enterococcus faecium and Enterococcus durans isolated from cheese: survival in the presence of medications under simulated gastrointestinal conditions and adhesion properties. J Dairy Sci. 2017;100(2):933–949. doi:10.3168/jds.2016-11513
21. Bhardwaj A, Gupta H, Kapila S, Kaur G, Vij S, Malik RK. Safety assessment and evaluation of probiotic potential of bacteriocinogenic Enterococcus faecium KH 24 strain under in vitro and in vivo conditions. Int J Food Microbiol. 2010;141(3):156–164. doi:10.1016/j.ijfoodmicro.2010.05.001
22. Guerrero-Ramos E, Cordero J, Molina-González D, et al. Antimicrobial resistance and virulence genes in enterococci from wild game meat in Spain. Food Microbiol. 2016;53:156–164. doi:10.1016/j.fm.2015.09.007
23. Byappanahalli MN, Nevers MB, Korajkic A, Staley ZR, Harwood VJ. Enterococci in the environment. Microbiol Mol Biol Rev. 2012;76(4):685–706. doi:10.1128/MMBR.00023-12
24. Seishima J, Iida N, Kitamura K, et al. Gut-derived Enterococcus faecium from ulcerative colitis patients promotes colitis in a genetically susceptible mouse host. Genome Biol. 2019;20:1–18. doi:10.1186/s13059-019-1879-9
25. Hayes JR, English LL, Carr LE, Wagner DD, Joseph SW. Multiple-antibiotic resistance of Enterococcus spp. isolated from commercial poultry production environments. Appl Environ Microbiol. 2004;70(10):6005–6011. doi:10.1128/AEM.70.10.6005-6011.2004
26. Rossitto P, Ruiz L, Kikuchi Y, et al. Antibiotic susceptibility patterns for environmental streptococci isolated from bovine mastitis in central California dairies. J Dairy Sci. 2002;85(1):132–138. doi:10.3168/jds.S0022-0302(02)74061-7
27. Tenhagen B-A, Köster G, Wallmann J, Heuwieser W. Prevalence of mastitis pathogens and their resistance against antimicrobial agents in dairy cows in Brandenburg, Germany. J Dairy Sci. 2006;89(7):2542–2551. doi:10.3168/jds.S0022-0302(06)72330-X
28. Ranasinghe R, Deshapriya R, Abeygunawardana D, Rahularaj R, Dematawewa C. Subclinical mastitis in dairy cows in major milk-producing areas of Sri Lanka: prevalence, associated risk factors, and effects on reproduction. J Dairy Sci. 2021;104(12):12900–12911. doi:10.3168/jds.2021-20223
29. Nam H, Lim S, Moon J, et al. Antimicrobial resistance of enterococci isolated from mastitic bovine milk samples in Korea. Zoonoses Public Health. 2010;57(7‐8):e59–e64. doi:10.1111/j.1863-2378.2009.01307.x
30. Yılmaz EŞ, Aslantaş Ö, Önen SP, Türkyılmaz S, Kürekci C. Prevalence, antimicrobial resistance and virulence traits in enterococci from food of animal origin in Turkey. LWT Food Sci Technol. 2016;66:20–26. doi:10.1016/j.lwt.2015.10.009
31. Esposito E, Campolo M, Casili G, et al. Protective effects of xyloglucan in association with the polysaccharide gelose in an experimental model of gastroenteritis and urinary tract infections. Int J Mol Sci. 2018;19(7):1844. doi:10.3390/ijms19071844
32. Piccinini D, Bernasconi E, Di Benedetto C, Martinetti Lucchini G, Bongiovanni M. Enterococcus hirae infections in the clinical practice. Infect Dis. 2023;55(1):71–73. doi:10.1080/23744235.2022.2125066
33. Gruet P, Maincent P, Berthelot X, Kaltsatos V. Bovine mastitis and intramammary drug delivery: review and perspectives. Adv Drug Deliv Rev. 2001;50(3):245–259. doi:10.1016/S0169-409X(01)00160-0
34. Vanderhaeghen W, Cerpentier T, Adriaensen C, Vicca J, Hermans K, Butaye P. Methicillin-resistant Staphylococcus aureus (MRSA) ST398 associated with clinical and subclinical mastitis in Belgian cows. Vet Microbiol. 2010;144(1–2):166–171. doi:10.1016/j.vetmic.2009.12.044
35. Friedman ND, Temkin E, Carmeli Y. The negative impact of antibiotic resistance. Clin Microbiol Infect. 2016;22(5):416–422. doi:10.1016/j.cmi.2015.12.002
36. Fiedler S, Bender J, Klare I, et al. Tigecycline resistance in clinical isolates of Enterococcus faecium is mediated by an upregulation of plasmid-encoded tetracycline determinants tet (L) and tet (M). J Antimicrob Chemother. 2016;71(4):871–881. doi:10.1093/jac/dkv420
37. Agga GE, Silva PJ, Martin RS. Tetracycline-and macrolide-resistant enterococcus species isolated from a Mink Farm in the United States. Microb Drug Resist. 2022;28(6):734–743. doi:10.1089/mdr.2021.0438
38. Ben Said L, Klibi N, Dziri R, et al. Prevalence, antimicrobial resistance and genetic lineages of Enterococcus spp. from vegetable food, soil and irrigation water in farm environments in Tunisia. J Sci Food Agric. 2016;96(5):1627–1633. doi:10.1002/jsfa.7264
39. Gazzola S, Fontana C, Bassi D, Cocconcelli P. Assessment of tetracycline and erythromycin resistance transfer during sausage fermentation by culture-dependent and-independent methods. Food Microbiol. 2012;30(2):348–354. doi:10.1016/j.fm.2011.12.005
40. Kaczorek E, Małaczewska J, Wójcik R, Rękawek W, Siwicki A. Phenotypic and genotypic antimicrobial susceptibility pattern of Streptococcus spp. isolated from cases of clinical mastitis in dairy cattle in Poland. J Dairy Sci. 2017;100(8):6442–6453. doi:10.3168/jds.2017-12660
41. Huys G, D’Haene K, Collard J-M, Swings J. Prevalence and molecular characterization of tetracycline resistance in Enterococcus isolates from food. Appl Environ Microbiol. 2004;70(3):1555–1562. doi:10.1128/AEM.70.3.1555-1562.2004
42. Arias CA, Contreras GA, Murray BE. Management of multidrug-resistant enterococcal infections. Clin Microbiol Infect. 2010;16(6):555–562. doi:10.1111/j.1469-0691.2010.03214.x
43. Lebreton F, van Schaik W, Manson McGuire A, et al. Emergence of epidemic multidrug-resistant Enterococcus faecium from animal and commensal strains. MBio. 2013;4(4):e00534–13. doi:10.1128/mBio.00534-13
44. Hegstad K, Mikalsen T, Coque TM, Werner G, Sundsfjord A. Mobile genetic elements and their contribution to the emergence of antimicrobial resistant Enterococcus faecalis and Enterococcus faecium. Clin Microbiol Infect. 2010;16(6):541–554. doi:10.1111/j.1469-0691.2010.03226.x
45. Manson AL, Van Tyne D, Straub TJ, et al. Chicken meat-associated enterococci: influence of agricultural antibiotic use and connection to the clinic. Appl Environ Microbiol. 2019;85(22):e01559–19. doi:10.1128/AEM.01559-19
46. Chajęcka-Wierzchowska W, Zarzecka U, Zadernowska A. Enterococci isolated from plant-derived food-analysis of antibiotic resistance and the occurrence of resistance genes. LWT. 2021;139:110549. doi:10.1016/j.lwt.2020.110549
47. Pezzella C, Ricci A, DiGiannatale E, Luzzi I, Carattoli A. Tetracycline and streptomycin resistance genes, transposons, and plasmids in Salmonella enterica isolates from animals in Italy. Antimicrob Agents Chemother. 2004;48(3):903–908. doi:10.1128/AAC.48.3.903-908.2004
48. Bolinger H, Kathariou S, Schaffner DW. The current state of macrolide resistance in Campylobacter spp.: trends and impacts of resistance mechanisms. Appl Environ Microbiol. 2017;83(12):e00416–17. doi:10.1128/AEM.00416-17
49. Chen W, Huang Y, Jiao X, Ye J, Lin Y, Yao F. Loss of erm (B)-mediated rRNA dimethylation and restoration of erythromycin susceptibility in erythromycin-resistant enterococci following induced linezolid resistance. Microb Drug Resist. 2023;29(2):51–58. doi:10.1089/mdr.2022.0140
50. Liu M, Douthwaite S. Activity of the ketolide telithromycin is refractory to Erm monomethylation of bacterial rRNA. Antimicrob Agents Chemother. 2002;46(6):1629–1633. doi:10.1128/AAC.46.6.1629-1633.2002
51. Brenciani A, Bacciaglia A, Vecchi M, Vitali LA, Varaldo PE, Giovanetti E. Genetic elements carrying erm (B) in Streptococcus pyogenes and association with tet (M) tetracycline resistance gene. Antimicrob Agents Chemother. 2007;51(4):1209–1216. doi:10.1128/AAC.01484-06
52. Schwaiger K, Harms K, Hölzel C, Meyer K, Karl M, Bauer J. Tetracycline in liquid manure selects for co-occurrence of the resistance genes tet (M) and tet (L) in Enterococcus faecalis. Vet Microbiol. 2009;139(3–4):386–392. doi:10.1016/j.vetmic.2009.06.005
53. Channaiah LH, Subramanyam B, Zurek L. Molecular characterization of antibiotic resistant and potentially virulent enterococci isolated from swine farms and feed mills. J Stored Prod Res. 2018;77:189–196. doi:10.1016/j.jspr.2018.04.007
54. Hammad AM, Shimamoto T, Shimamoto T. Genetic characterization of antibiotic resistance and virulence factors in Enterococcus spp. from Japanese retail ready-to-eat raw fish. Food Microbiol. 2014;38:62–66. doi:10.1016/j.fm.2013.08.010
55. Singh KV, Nallapareddy SR, Sillanpää J, Murray BE. Importance of the collagen adhesin ace in pathogenesis and protection against Enterococcus faecalis experimental endocarditis. PLoS Pathogens. 2010;6(1):e1000716. doi:10.1371/annotation/1ccae8f8-d274-4ff8-a295-815037ce9cc6
56. Stępień-Pyśniak D, Hauschild T, Kosikowska U, Dec M, Urban-Chmiel R. Biofilm formation capacity and presence of virulence factors among commensal Enterococcus spp. from wild birds. Sci Rep. 2019;9(1):1–7. doi:10.1038/s41598-019-47602-w
57. Martín I, Barbosa J, Pereira SI, Rodríguez A, Córdoba JJ, Teixeira P. Study of lactic acid bacteria isolated from traditional ripened foods and partial characterization of their bacteriocins. LWT. 2023;173:114300. doi:10.1016/j.lwt.2022.114300
58. Kongsted H, Pedersen K, Hjulsager CK, et al. Diarrhoea in neonatal piglets: a case control study on microbiological findings. Porcine Health Manage. 2018;4(1):1–7. doi:10.1186/s40813-018-0094-5
59. Oruc O, Cetin O, Darilmaz DO, Yüsekdag ZN. Determination of the biosafety of potential probiotic Enterococcus faecalis and Enterococcus faecium strains isolated from traditional white cheeses. LWT. 2021;148:111741. doi:10.1016/j.lwt.2021.111741
60. Soares RO, Fedi AC, Reiter KC, Caierão J, d’Azevedo PA. Correlation between biofilm formation and gelE, esp, and agg genes in Enterococcus spp. clinical isolates. Virulence. 2014;5(5):634–637. doi:10.4161/viru.28998
61. Chajęcka-Wierzchowska W, Zadernowska A, Łaniewska-Trokenheim Ł. Virulence factors, antimicrobial resistance and biofilm formation in Enterococcus spp. isolated from retail shrimps. LWT Food Sci Technol. 2016;69:117–122. doi:10.1016/j.lwt.2016.01.034
62. Franz CM, Huch M, Abriouel H, Holzapfel W, Gálvez A. Enterococci as probiotics and their implications in food safety. Int J Food Microbiol. 2011;151(2):125–140. doi:10.1016/j.ijfoodmicro.2011.08.014
63. El‐Ghaish S, Khalifa M, Elmahdy A. Antimicrobial impact for Lactococcus lactis subsp. lactis A15 and Enterococcus faecium A15 isolated from some traditional Egyptian dairy products on some pathogenic bacteria. J Food Biochem. 2017;41(1):e12279. doi:10.1111/jfbc.12279
64. Bennett RJ, Dunny GM. Analogous telesensing pathways regulate mating and virulence in two opportunistic human pathogens. Mbio. 2010;1(4):e00181–10. doi:10.1128/mBio.00181-10
65. Dunny GM. Enterococcal sex pheromones: signaling, social behavior, and evolution. Ann Rev Genet. 2013;47(1):457–482. doi:10.1146/annurev-genet-111212-133449
66. Duan Y, Llorente C, Lang S, et al. Bacteriophage targeting of gut bacterium attenuates alcoholic liver disease. Nature. 2019;575(7783):505–511. doi:10.1038/s41586-019-1742-x
67. Jett B, Jensen H, Nordquist R, Gilmore M. Contribution of the pAD1-encoded cytolysin to the severity of experimental Enterococcus faecalis endophthalmitis. Infect Immun. 1992;60(6):2445–2452. doi:10.1128/iai.60.6.2445-2452.1992
68. Franz CM, Muscholl-Silberhorn AB, Yousif NM, Vancanneyt M, Swings J, Holzapfel WH. Incidence of virulence factors and antibiotic resistance among enterococci isolated from food. Appl Environ Microbiol. 2001;67(9):4385–4389. doi:10.1128/AEM.67.9.4385-4389.2001
69. Mundy L, Sahm D, Gilmore M. Relationships between enterococcal virulence and antimicrobial resistance. Clin Microbiol Rev. 2000;13(4):513–522. doi:10.1128/CMR.13.4.513
70. Hammad AM, Hassan HA, Shimamoto T. Prevalence, antibiotic resistance and virulence of Enterococcus spp. in Egyptian fresh raw milk cheese. Food Control. 2015;50:815–820. doi:10.1016/j.foodcont.2014.10.020
71. Semedo T, Almeida Santos M, Martins P, et al. Comparative study using type strains and clinical and food isolates to examine hemolytic activity and occurrence of the cyl operon in enterococci. J Clin Microbiol. 2003;41(6):2569–2576. doi:10.1128/JCM.41.6.2569-2576.2003
72. Bebien M, Hensler ME, Davanture S, et al. The pore-forming toxin β hemolysin/cytolysin triggers p38 MAPK-dependent IL-10 production in macrophages and inhibits innate immunity. PLoS Pathogens. 2012;8(7):e1002812. doi:10.1371/journal.ppat.1002812
73. Vankerckhoven V, Van Autgaerden T, Vael C, et al. Development of a multiplex PCR for the detection of asa1, gelE, cylA, esp, and hyl genes in enterococci and survey for virulence determinants among European hospital isolates of Enterococcus faecium. J Clin Microbiol. 2004;42(10):4473–4479. doi:10.1128/JCM.42.10.4473-4479.2004
74. Shankar V, Baghdayan AS, Huycke MM, Lindahl G, Gilmore MS, Tuomanen EI. Infection-derived Enterococcus faecalis strains are Enriched in esp, a gene encoding a novel surface protein. Infect Immun. 1999;67(1):193–200. doi:10.1128/IAI.67.1.193-200.1999
75. Gholizadeh P, Aghazadeh M, Ghotaslou R, et al. CRISPR-cas system in the acquisition of virulence genes in dental-root canal and hospital-acquired isolates of Enterococcus faecalis. Virulence. 2020;11(1):1257–1267. doi:10.1080/21505594.2020.1809329
76. Arciola CR, Baldassarri L, Campoccia D, et al. Strong biofilm production, antibiotic multi-resistance and high gelE expression in epidemic clones of Enterococcus faecalis from orthopaedic implant infections. Biomaterials. 2008;29(5):580–586. doi:10.1016/j.biomaterials.2007.10.008
77. Sifri CD, Mylonakis E, Singh KV, et al. Virulence effect of Enterococcus faecalis protease genes and the quorum-sensing locus fsr in Caenorhabditis elegans and mice. Infect Immun. 2002;70(10):5647–5650. doi:10.1128/IAI.70.10.5647-5650.2002
 © 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.